You might also like
- Aula Parametros Genéticos PDFDocument20 pagesAula Parametros Genéticos PDFKaio FernandesNo ratings yet
- Heterose e HeterozigoseDocument25 pagesHeterose e Heterozigoselucildalima100% (1)
- Soro Da Memória Do Leite - Globo Reporter - Rede GloboDocument2 pagesSoro Da Memória Do Leite - Globo Reporter - Rede GloboAndré SilverioNo ratings yet
- Anatomia Do Aparelho Reprodutor Do MachoDocument26 pagesAnatomia Do Aparelho Reprodutor Do MachoAlexandre Silva de AlmeidaNo ratings yet
- Pelagem e Raças Dos EquinosDocument75 pagesPelagem e Raças Dos Equinosmilena_airesNo ratings yet
- Peritonite em EquinosDocument16 pagesPeritonite em EquinosDaizy QueirozNo ratings yet
- AULA 03 - Fermentação RuminalDocument28 pagesAULA 03 - Fermentação RuminalFlávio Moreno100% (1)
- Lipidios Na Nutricao de RuminantesDocument14 pagesLipidios Na Nutricao de Ruminantesamamùra maamarNo ratings yet
- Cloro e Molibdênio Nutrição AnimalDocument9 pagesCloro e Molibdênio Nutrição AnimalGeórgia BonifácioNo ratings yet
- Anatomia Topográfica Do Casco EquinoDocument6 pagesAnatomia Topográfica Do Casco EquinoAlexandro SilvaNo ratings yet
- 8-Exame Andrológico Do TouroDocument52 pages8-Exame Andrológico Do Tourolucaskkk123No ratings yet
- Causalidade em EpidemiologiaDocument33 pagesCausalidade em EpidemiologiaPedro HenriqueNo ratings yet
- Melhoramento Genetico em Gado LeiteiroDocument41 pagesMelhoramento Genetico em Gado LeiteiroDelano AlencarNo ratings yet
- Manual de Exame Ginecológico Da VacaDocument5 pagesManual de Exame Ginecológico Da VacamarciogomesgvNo ratings yet
- Metabolismo e Digestão de Lipídios em Vacas LeiteirasDocument43 pagesMetabolismo e Digestão de Lipídios em Vacas Leiteirascedapsan100% (1)
- Bovinos de Corte CruzamentosDocument83 pagesBovinos de Corte Cruzamentosa097792No ratings yet
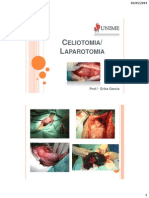
- Celioto MiaDocument20 pagesCelioto MiaLuiza NavarroNo ratings yet
- CruzamentoDocument11 pagesCruzamentoVet_arquivos100% (2)
- Digestão e Absorção de Produtos 2020.2Document61 pagesDigestão e Absorção de Produtos 2020.2Tata PetroffNo ratings yet
- Métodos Semiológicos de Exame Físico em RuminantesDocument5 pagesMétodos Semiológicos de Exame Físico em RuminantesVet_arquivos100% (3)
- Melhoramento Animal Aplicado I PDFDocument2 pagesMelhoramento Animal Aplicado I PDFplfs1974No ratings yet
- Aspectos Gerais Da Cirurgia AbdominalDocument17 pagesAspectos Gerais Da Cirurgia AbdominalMarcy AraujoNo ratings yet
- Bos Taurus Indicus (Zebuínos)Document33 pagesBos Taurus Indicus (Zebuínos)anelise_100% (1)
- Aula8 Desenho de PrimersDocument59 pagesAula8 Desenho de PrimersCarlianeNo ratings yet
- DTA - AulaDocument19 pagesDTA - AulaRodrigoNo ratings yet
- 1 - Introdução A AnatomiaDocument80 pages1 - Introdução A AnatomiaTaynara BarreiraNo ratings yet
- Exame Ginecológico em Pequeno AnimaisDocument7 pagesExame Ginecológico em Pequeno AnimaisGrazi HonorioNo ratings yet
- Nutrição Animal BásicaDocument36 pagesNutrição Animal BásicaLuana CristianeNo ratings yet
- Livro Professor Vidal Vol01 BAIXADocument123 pagesLivro Professor Vidal Vol01 BAIXAMarciel FrançaNo ratings yet
- Afecções Locomotoras Traumáticas em EqüinosDocument55 pagesAfecções Locomotoras Traumáticas em EqüinosLuiz Henrique Brandão PiresNo ratings yet
- Doce de LeiteDocument3 pagesDoce de LeiteÉrika PatriciaNo ratings yet
- Otite Média em Equino Decorrente de Intoxicação Por Graxa Automotiva: Relato de CasoDocument1 pageOtite Média em Equino Decorrente de Intoxicação Por Graxa Automotiva: Relato de CasoKarina CalNo ratings yet
- 1583408338GUIA TCNICO - Passos para o Sucesso Na Inseminao Artificial PDFDocument46 pages1583408338GUIA TCNICO - Passos para o Sucesso Na Inseminao Artificial PDFRubens BorgesNo ratings yet
- Aula 2 - Genética de Populações PDFDocument54 pagesAula 2 - Genética de Populações PDFClaudemir TrevisanNo ratings yet
- Diferencial de Seleção e Ganho GenéticoDocument14 pagesDiferencial de Seleção e Ganho Genéticolucildalima100% (5)
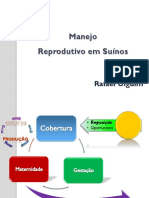
- Manejo Reprodutivo em SuinosDocument55 pagesManejo Reprodutivo em SuinosPedro SantosNo ratings yet
- Pododermatite Infecciosa em Equino Associada A Osteíte de Terceira Falange: Relato de CasoDocument1 pagePododermatite Infecciosa em Equino Associada A Osteíte de Terceira Falange: Relato de CasoKarina CalNo ratings yet
- 11 - Transferencia de Embrioes BovinosDocument75 pages11 - Transferencia de Embrioes BovinosMirela Cristine Pereira100% (1)
- Aula SarnasDocument59 pagesAula SarnasGedeao Candido CamisaNo ratings yet
- EXERCÍCIOS DE MELHORAMENTO - Graus de SangueDocument3 pagesEXERCÍCIOS DE MELHORAMENTO - Graus de SangueAmanda TramonteNo ratings yet
- Inseminação Artificial em SuinosDocument90 pagesInseminação Artificial em SuinosJúlio Cesar PereiraNo ratings yet
- ApostilaDocument30 pagesApostilaJorge ReppoldNo ratings yet
- DEP: Como Calcular e Como UtilizarDocument5 pagesDEP: Como Calcular e Como Utilizaravantidigital100% (2)
- Genética QuantitativaDocument59 pagesGenética Quantitativalucildalima100% (1)
- Aula 9 Lactação Fisio II 2020Document54 pagesAula 9 Lactação Fisio II 2020Betânia Glória CamposNo ratings yet
- OxyurideaDocument40 pagesOxyurideawellempormesNo ratings yet
- Aula 04 - Fisiologia e Endocrinologia Da Reprodução Da FêmeaDocument74 pagesAula 04 - Fisiologia e Endocrinologia Da Reprodução Da FêmeaHELIDA FERNANDES LEAONo ratings yet
- Equinocultura - ResenhaDocument31 pagesEquinocultura - ResenhaRaimundo Franco100% (1)
- Equívocos Na Cólica EquinaDocument7 pagesEquívocos Na Cólica EquinaRaíssa GalegoNo ratings yet
- Anuario Nutricao Vegetal Abisolo2015Document124 pagesAnuario Nutricao Vegetal Abisolo2015Núbia NunesNo ratings yet
- Manejo Nutricional de Vacas em LactaçãoDocument22 pagesManejo Nutricional de Vacas em Lactaçãomarllinh0No ratings yet
- Genoma BovinoDocument18 pagesGenoma BovinoFlávio LimaNo ratings yet
- Semen SexadoDocument22 pagesSemen Sexadomeyer66No ratings yet
- 2 - Fisiologia Do Ciclo Estral Nas Especies Domesticas GabrielDocument57 pages2 - Fisiologia Do Ciclo Estral Nas Especies Domesticas GabrielHériton SilvaNo ratings yet
- Palestra Parto e Distocias em Cães - Quando IntervirDocument28 pagesPalestra Parto e Distocias em Cães - Quando IntervirFernanda Trigo de AzevedoNo ratings yet
- Abscesso Hepático em Bovinos de AbateDocument12 pagesAbscesso Hepático em Bovinos de AbateFlavio alcantara silvaNo ratings yet
- 9 Índices Zootécnicos Recomendados de Acordo Com Cada EspécieDocument10 pages9 Índices Zootécnicos Recomendados de Acordo Com Cada EspécieMaria Ercilia FernandesNo ratings yet
- Controle Zootecnico em Bovinos de CorteDocument3 pagesControle Zootecnico em Bovinos de CorteGelson LodiNo ratings yet
- (PDF) Demodicose Canina PDFDocument15 pages(PDF) Demodicose Canina PDFVet_arquivosNo ratings yet
- Wasava 2015 - Diretrizes para A Vacinação de Cães e Gatos PDFDocument50 pagesWasava 2015 - Diretrizes para A Vacinação de Cães e Gatos PDFmarcelo100% (1)
- Dicionário Inglês-Português de SuinoculturaDocument30 pagesDicionário Inglês-Português de SuinoculturaLeandro CastilhaNo ratings yet
- Curso VendedorDocument138 pagesCurso VendedorVet_arquivosNo ratings yet
- Manual Pos VacinaçãoDocument188 pagesManual Pos VacinaçãoTamara Coutinho Galvão SilvaNo ratings yet
- Gênero Leptospira 2015Document52 pagesGênero Leptospira 2015Vet_arquivosNo ratings yet
- Bravecto DemodecicaDocument7 pagesBravecto DemodecicaVet_arquivosNo ratings yet
- Cartilha Animais PeçonhentosDocument24 pagesCartilha Animais PeçonhentosAlan DemarcosNo ratings yet
- As Prostaglandinas Na ReproduçãoDocument26 pagesAs Prostaglandinas Na ReproduçãoVet_arquivos100% (2)
- Sarna Sarcoptica em Caes PDFDocument5 pagesSarna Sarcoptica em Caes PDFMJoanaDuarteNo ratings yet
- Controle Endócrino Do Equilíbrio Hidro-EletrolíticoDocument18 pagesControle Endócrino Do Equilíbrio Hidro-EletrolíticoVet_arquivosNo ratings yet
- Timpanismo em RuminAntesDocument6 pagesTimpanismo em RuminAntesVet_arquivos100% (1)
- Choque Circulatorio-Aspectos Básicos de Fisiopatologia e TerapeuticaDocument22 pagesChoque Circulatorio-Aspectos Básicos de Fisiopatologia e TerapeuticaVet_arquivosNo ratings yet
- Qualidade Do Colostro BovinoDocument5 pagesQualidade Do Colostro BovinoVet_arquivosNo ratings yet
- Noções Básicas de FarmacologiaDocument16 pagesNoções Básicas de FarmacologiaVet_arquivos100% (5)
- Retenção de Placenta em VacasDocument7 pagesRetenção de Placenta em VacasVet_arquivos100% (1)
- Carbúnculo Sintomático em BovinoDocument5 pagesCarbúnculo Sintomático em BovinoVet_arquivosNo ratings yet
- Carbúnculo HemáticoDocument5 pagesCarbúnculo HemáticoVet_arquivos100% (1)
- SuinoculturaDocument27 pagesSuinoculturaVet_arquivos100% (3)
- Causa e Tratamento Do Garrotilho em EquinosDocument5 pagesCausa e Tratamento Do Garrotilho em EquinosVet_arquivos100% (1)
- Terapêutica de Aves SilvestresDocument6 pagesTerapêutica de Aves SilvestresVet_arquivosNo ratings yet
- A Importância Do Médico Veterinário Na Saúde PúblicaDocument47 pagesA Importância Do Médico Veterinário Na Saúde PúblicaVet_arquivos100% (3)
- Doenças Respiratórias Dos BovinosDocument6 pagesDoenças Respiratórias Dos BovinosVet_arquivosNo ratings yet
- Digestão de Celulose Nos VertebradosDocument2 pagesDigestão de Celulose Nos VertebradosVet_arquivosNo ratings yet
- Exame Clínico de Enfermidades Do Sist Nervoso em Equinos e RuminantesDocument17 pagesExame Clínico de Enfermidades Do Sist Nervoso em Equinos e RuminantesVet_arquivosNo ratings yet
- Métodos Semiológicos de Exame Físico em RuminantesDocument5 pagesMétodos Semiológicos de Exame Físico em RuminantesVet_arquivos100% (3)
- Sistema Digestivo Dos Rum in AntesDocument6 pagesSistema Digestivo Dos Rum in AntesVet_arquivosNo ratings yet
- Raças de CãesDocument98 pagesRaças de CãesVet_arquivos100% (4)
- Alguns Pontos Importantes em Exames Clínicos de AnimaisDocument4 pagesAlguns Pontos Importantes em Exames Clínicos de AnimaisVet_arquivosNo ratings yet
- Inovações Técnológicas Na BioclimatologiaDocument24 pagesInovações Técnológicas Na BioclimatologiaVet_arquivos100% (1)
- Mapeamento... Ebook IfsertãoDocument260 pagesMapeamento... Ebook IfsertãoAdelson Dias OliveiraNo ratings yet
- Plano Analitco Geografia 8 Classe - I Trimestre 2024Document4 pagesPlano Analitco Geografia 8 Classe - I Trimestre 2024Melshim Marcelo LopesNo ratings yet
- Escala 2023 - CratoDocument13 pagesEscala 2023 - CratocristinaraimundoNo ratings yet
- Grandes Temas em Biologia: MódulosDocument255 pagesGrandes Temas em Biologia: Móduloseduardo100% (1)
- Plano Diretor VG 1 PDFDocument174 pagesPlano Diretor VG 1 PDFCezar ClementeNo ratings yet
- EVOLUCAODocument13 pagesEVOLUCAOSelemane AtijaNo ratings yet
- Exames de Geografia CompiladosDocument363 pagesExames de Geografia CompiladosAndréXavierNo ratings yet
- Aulas - 5 AnoDocument3 pagesAulas - 5 AnoJéssica Lima LopesNo ratings yet
- DocumentoDocument7 pagesDocumentoSylvyaNo ratings yet
- Prova - PB .Linguaportuguesa.4ano - Manha .3bimDocument5 pagesProva - PB .Linguaportuguesa.4ano - Manha .3bimSilvania Pires De Oliveira AraujoNo ratings yet
- Geo20 08Document3 pagesGeo20 08Ibrahim AlhannaNo ratings yet
- 5 Ecologia de MetapopulaçãoDocument31 pages5 Ecologia de MetapopulaçãoMarcos AurelioNo ratings yet
- Herdabilidade e RepetibilidadeDocument14 pagesHerdabilidade e RepetibilidadeAlexandre AssisNo ratings yet
- Cartilha Povos OrigináriosDocument6 pagesCartilha Povos OrigináriosMayara HelenaNo ratings yet
- Exame de Geografia55Document5 pagesExame de Geografia55vivaldoNo ratings yet
- PDF c16 001 016 VDG SP Aula p02 m17Document16 pagesPDF c16 001 016 VDG SP Aula p02 m17Lucas Biotto MachadoNo ratings yet
- Evolução Da PopulaçãoDocument3 pagesEvolução Da PopulaçãoVictóriaNo ratings yet
- 2º Trimestre Avaliação de Geografia SobreDocument4 pages2º Trimestre Avaliação de Geografia SobreZaria Gonçalves DA SilvaNo ratings yet
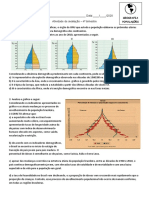
- O Crescimento Da População Mundial Até 2100Document4 pagesO Crescimento Da População Mundial Até 2100José Eustáquio Diniz AlvesNo ratings yet
- Pirâmide Etária - AtividadesDocument2 pagesPirâmide Etária - AtividadesmiltinoNo ratings yet
- SimuunivertixDocument5 pagesSimuunivertixigor25% (4)
- 7 Ano 3 BimestreDocument5 pages7 Ano 3 BimestreJhoniesley Carvalho de OliveiraNo ratings yet
- Residuos Solidos em Imperatriz MADocument309 pagesResiduos Solidos em Imperatriz MAMargarida da ConceiçãoNo ratings yet
- Gestão Da Drenagem Urbana Na Bacia Hidrográfica Do Rio Itajaí AçuDocument142 pagesGestão Da Drenagem Urbana Na Bacia Hidrográfica Do Rio Itajaí AçuFabitascaNo ratings yet
- Lista de Vacinados de Dezembro de 2021Document10 pagesLista de Vacinados de Dezembro de 2021luiz kawaiNo ratings yet
- Data de Entredas ProjetosDocument1 pageData de Entredas Projetoseuzielly.tomazNo ratings yet
- Historico ItaqueraDocument4 pagesHistorico ItaqueraCaio Cobucci LeiteNo ratings yet
- CC Bueno 00779581016Document9 pagesCC Bueno 00779581016FernandaNo ratings yet
- Regiões BrasileirasDocument5 pagesRegiões BrasileirasDaviGomideNo ratings yet
- Aula 03 - Genética Quantitativa - Poligenes, Herdabilidade, Ganho de Seleção e Heterose - Sem RespostaDocument67 pagesAula 03 - Genética Quantitativa - Poligenes, Herdabilidade, Ganho de Seleção e Heterose - Sem RespostarakelNo ratings yet