You might also like
- Congenital Glaucomas: 1. Primary 2. Iridocorneal DysgenesisDocument12 pagesCongenital Glaucomas: 1. Primary 2. Iridocorneal DysgenesiskhadzxNo ratings yet
- Dermatologic DiseasesDocument2 pagesDermatologic DiseaseskhadzxNo ratings yet
- 34 Idiopathic Spec Uveitis SyndromeDocument8 pages34 Idiopathic Spec Uveitis SyndromekhadzxNo ratings yet
- 6th Central Pay Commission Salary CalculatorDocument15 pages6th Central Pay Commission Salary Calculatorrakhonde100% (436)
- Metabolic Bone DiseaseDocument29 pagesMetabolic Bone DiseasekhadzxNo ratings yet
- 13 Peripheral Corneal InflamDocument5 pages13 Peripheral Corneal InflamkhadzxNo ratings yet
- Acquired 2. Isolated Familial Ectopia Lentis 3. Associated With Systemic SyndromesDocument8 pagesAcquired 2. Isolated Familial Ectopia Lentis 3. Associated With Systemic SyndromeskhadzxNo ratings yet
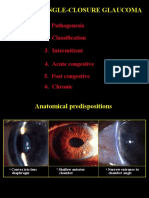
- Primary Angle-Closure Glaucoma: 1. Pathogenesis 2. ClassificationDocument9 pagesPrimary Angle-Closure Glaucoma: 1. Pathogenesis 2. ClassificationkhadzxNo ratings yet
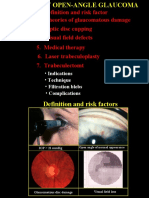
- 28primary Open Angle GlaucomaDocument21 pages28primary Open Angle GlaucomakhadzxNo ratings yet
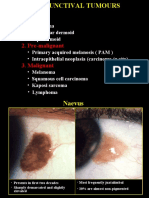
- 12conjunctival TumoursDocument12 pages12conjunctival TumourskhadzxNo ratings yet
- Congenital Cataract: 1. Important FactsDocument13 pagesCongenital Cataract: 1. Important FactskhadzxNo ratings yet
- Juvenile Rheumatoid Arthritis (Jra) (Juvenile Chronic Arthritis-Jca)Document70 pagesJuvenile Rheumatoid Arthritis (Jra) (Juvenile Chronic Arthritis-Jca)khadzxNo ratings yet
- Abnormal Lens ShapeDocument4 pagesAbnormal Lens ShapekhadzxNo ratings yet
- 15corneal InfectionsDocument8 pages15corneal InfectionskhadzxNo ratings yet
- 09conjunctival InfectionsDocument11 pages09conjunctival InfectionskhadzxNo ratings yet
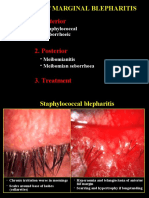
- 02chronic Marginal BlepharitisDocument7 pages02chronic Marginal BlepharitiskhadzxNo ratings yet
- Ndiang'Ui: Unexpected Deaths 24 September 2009Document25 pagesNdiang'Ui: Unexpected Deaths 24 September 2009khadzxNo ratings yet
- Disorders of Eye LashesDocument8 pagesDisorders of Eye LasheskhadzxNo ratings yet
- Poisoning by MedicinesDocument11 pagesPoisoning by MedicineskhadzxNo ratings yet
- Postmortem Changes and Postmortem Time IntervalDocument24 pagesPostmortem Changes and Postmortem Time IntervalkhadzxNo ratings yet
- Chronic Osteomyelitis in Early Infancy: Presenter: DR Maina Discussant: DR Mogire (Orthopedic Surgeon)Document27 pagesChronic Osteomyelitis in Early Infancy: Presenter: DR Maina Discussant: DR Mogire (Orthopedic Surgeon)khadzxNo ratings yet
- External Fixation of The Lower Limb 2Document13 pagesExternal Fixation of The Lower Limb 2khadzxNo ratings yet
- Sudden Unexpected Death FMT 400Document11 pagesSudden Unexpected Death FMT 400khadzxNo ratings yet
- Narcotics N HallucinogensDocument14 pagesNarcotics N HallucinogenskhadzxNo ratings yet
- Introduction To Forensic Medicine DR David Chumba MBCHB, Mmed (Human Pathology), Dip. For Med (Sa)Document20 pagesIntroduction To Forensic Medicine DR David Chumba MBCHB, Mmed (Human Pathology), Dip. For Med (Sa)khadzxNo ratings yet
- Introduction To Forensic Medicine DR David Chumba MBCHB, Mmed (Human Pathology), Dip. For Med (Sa)Document20 pagesIntroduction To Forensic Medicine DR David Chumba MBCHB, Mmed (Human Pathology), Dip. For Med (Sa)khadzxNo ratings yet
- Fire Arm Injuries 4th Yr ForensicDocument31 pagesFire Arm Injuries 4th Yr ForensickhadzxNo ratings yet
- Carbon Monoxide Poisoning 4th YrDocument17 pagesCarbon Monoxide Poisoning 4th YrkhadzxNo ratings yet
- Classification of Injuries FMTDocument30 pagesClassification of Injuries FMTkhadzx100% (2)
- Malocclusion and Orthodontic Eldoret 2009Document33 pagesMalocclusion and Orthodontic Eldoret 2009khadzxNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- 10CV54 Unit 05 PDFDocument21 pages10CV54 Unit 05 PDFvinodh159No ratings yet
- Project Final Report: Crop BreedingDocument16 pagesProject Final Report: Crop BreedingAniket PatilNo ratings yet
- Ds 3805Document4 pagesDs 3805sparky2017No ratings yet
- Citation GuideDocument21 pagesCitation Guideapi-229102420No ratings yet
- Amex Case StudyDocument12 pagesAmex Case StudyNitesh JainNo ratings yet
- Talon Star Trek Mod v0.2Document4 pagesTalon Star Trek Mod v0.2EdmundBlackadderIVNo ratings yet
- Robin Engine EH722 DS 7010Document29 pagesRobin Engine EH722 DS 7010yewlimNo ratings yet
- Performance of a Pelton WheelDocument17 pagesPerformance of a Pelton Wheellimakupang_matNo ratings yet
- What Is Inventory Management?Document31 pagesWhat Is Inventory Management?Naina SobtiNo ratings yet
- ROM Magazine V1i6Document64 pagesROM Magazine V1i6Mao AriasNo ratings yet
- 1 20《经济学家》读译参考Document62 pages1 20《经济学家》读译参考xinying94No ratings yet
- (V) 2020-Using Extensive Reading in Improving Reading Speed and Level of Reading Comprehension of StudentsDocument7 pages(V) 2020-Using Extensive Reading in Improving Reading Speed and Level of Reading Comprehension of StudentsMEYTA RAHMATUL AZKIYANo ratings yet
- Plumbing Arithmetic RefresherDocument80 pagesPlumbing Arithmetic RefresherGigi AguasNo ratings yet
- Where Are The Women in The Water Pipeline? Wading Out of The Shallows - Women and Water Leadership in GeorgiaDocument7 pagesWhere Are The Women in The Water Pipeline? Wading Out of The Shallows - Women and Water Leadership in GeorgiaADBGADNo ratings yet
- Irc SP 65-2005 PDFDocument32 pagesIrc SP 65-2005 PDFAjay Kumar JainNo ratings yet
- Chapter 1Document2 pagesChapter 1Nor-man KusainNo ratings yet
- BILL of Entry (O&A) PDFDocument3 pagesBILL of Entry (O&A) PDFHiJackNo ratings yet
- 4th Summative Test Science 6Document5 pages4th Summative Test Science 6ANNALIZA FIECASNo ratings yet
- Lignan & NeolignanDocument12 pagesLignan & NeolignanUle UleNo ratings yet
- Module 7 - Assessment of Learning 1 CoursepackDocument7 pagesModule 7 - Assessment of Learning 1 CoursepackZel FerrelNo ratings yet
- Working With Session ParametersDocument10 pagesWorking With Session ParametersyprajuNo ratings yet
- Monitoring Tool in ScienceDocument10 pagesMonitoring Tool in ScienceCatherine RenanteNo ratings yet
- Case 1 1 Starbucks Going Global FastDocument2 pagesCase 1 1 Starbucks Going Global FastBoycie TarcaNo ratings yet
- Minimum Fees To Be Taken by CADocument8 pagesMinimum Fees To Be Taken by CACA Sanjay BhatiaNo ratings yet
- Galley cleaning hazards and controlsDocument9 pagesGalley cleaning hazards and controlsRosalie RosalesNo ratings yet
- Windows Keyboard Shortcuts OverviewDocument3 pagesWindows Keyboard Shortcuts OverviewShaik Arif100% (1)
- BWT Septron Line 31-61 Rev01!08!05-18 Opm enDocument56 pagesBWT Septron Line 31-61 Rev01!08!05-18 Opm enDavide Grioni100% (1)
- 2 Case StudyDocument8 pages2 Case Studysehrish khawerNo ratings yet
- PSAII Final EXAMDocument15 pagesPSAII Final EXAMdaveadeNo ratings yet
- Corporate GovernanceDocument35 pagesCorporate GovernanceshrikirajNo ratings yet