You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5784)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (72)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Function of RBCDocument9 pagesFunction of RBCAthea MelosantosNo ratings yet
- Preservation and Conservation of EnvironmentDocument3 pagesPreservation and Conservation of EnvironmentCloe Dianne SillaNo ratings yet
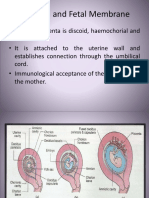
- Placenta and Fetal Membrane-6614Document45 pagesPlacenta and Fetal Membrane-6614Incredible DivineNo ratings yet
- General Pathology (Lecture-1) - 3Document14 pagesGeneral Pathology (Lecture-1) - 3Usman AkramNo ratings yet
- 2686 PDFDocument11 pages2686 PDFmojNo ratings yet
- Name Class DateDocument2 pagesName Class Dateapi-25965241No ratings yet
- Understanding Apicomplexan ParasitesDocument51 pagesUnderstanding Apicomplexan ParasitesruthNo ratings yet
- Cladogram CH 19 Part ADocument3 pagesCladogram CH 19 Part AJasmine lee100% (1)
- First Year B.Sc. Nursing Question Paper 1998Document3 pagesFirst Year B.Sc. Nursing Question Paper 1998Biju AntonyNo ratings yet
- ABi 7300 BrochureDocument4 pagesABi 7300 BrochureAndry WigunaNo ratings yet
- ISC 11th - Biology 2023-24 Council Syllabus-1-10Document10 pagesISC 11th - Biology 2023-24 Council Syllabus-1-10R HarryNo ratings yet
- PROTOZOA DARAH (MALARIADocument88 pagesPROTOZOA DARAH (MALARIAGladis Aprilla RizkiNo ratings yet
- A Checklist of Weevils of The Tribe Mecysolobini (Coleoptera Curculionidae Molytinae) PDFDocument67 pagesA Checklist of Weevils of The Tribe Mecysolobini (Coleoptera Curculionidae Molytinae) PDFNick Anthony BuriasNo ratings yet
- Science Technology and Society Chapter 3Document76 pagesScience Technology and Society Chapter 3JOHN RONALD SAJOL67% (3)
- PdcaasDocument8 pagesPdcaasTan Suk WanNo ratings yet
- Gender Is It A Social Construct or A Biological InevitabilityDocument5 pagesGender Is It A Social Construct or A Biological InevitabilitySaurabhChoudharyNo ratings yet
- 1989 - Alterations of Life Span in The Nematode Caenorhabditis Elegans Under Monoxenic Culture ConditionsDocument14 pages1989 - Alterations of Life Span in The Nematode Caenorhabditis Elegans Under Monoxenic Culture ConditionsRaymond LaBoyNo ratings yet
- Walk-In-Interview in CRPFDocument1 pageWalk-In-Interview in CRPFDevi PrasadNo ratings yet
- Plant Anatomy: Structure and FunctionsDocument8 pagesPlant Anatomy: Structure and FunctionsWimartama WayanNo ratings yet
- Drug DesignDocument38 pagesDrug DesignPhArMaCyGrAdUaTeSNo ratings yet
- Osmolarity&tonicitypracticeproblemsfrominternetDocument14 pagesOsmolarity&tonicitypracticeproblemsfrominternetbsktblbabe88No ratings yet
- Parallel Evolution of Cannabinoid Biosynthesis: Nature PlantsDocument31 pagesParallel Evolution of Cannabinoid Biosynthesis: Nature PlantsPaulo EdsonNo ratings yet
- Bryophyta Water Relations PDFDocument40 pagesBryophyta Water Relations PDFCDB 1st Semester 2077No ratings yet
- UGCNET Paper2 AnthropologyDocument56 pagesUGCNET Paper2 AnthropologyAryanNo ratings yet
- 3.2. Keanekaragaman Makhluk Hidup (Fungi)Document42 pages3.2. Keanekaragaman Makhluk Hidup (Fungi)Bagus Dwi PermanaNo ratings yet
- Chapter 5 Evolution and Biodiversity Exercises IB SLDocument8 pagesChapter 5 Evolution and Biodiversity Exercises IB SLSanaa SamkoNo ratings yet
- Chapter 03Document4 pagesChapter 03Teehee JonesNo ratings yet
- January 2012 QP - Unit 4 Edexcel Biology A-LevelDocument24 pagesJanuary 2012 QP - Unit 4 Edexcel Biology A-LevelElissa MeflehNo ratings yet
- Biology Module 1 Lesson 3 Recombinant DNA TechnologyDocument15 pagesBiology Module 1 Lesson 3 Recombinant DNA TechnologyHannah Joy D. AROSTIQUENo ratings yet
- Revision For Mid Term Test 2.: Module: C - Importance of PlantsDocument10 pagesRevision For Mid Term Test 2.: Module: C - Importance of Plantsvenice tranNo ratings yet