You might also like
- Razonamiento lógico primariaDocument80 pagesRazonamiento lógico primariaMartitaossi OssorioNo ratings yet
- Cuidados Del Recién NacidoDocument18 pagesCuidados Del Recién NacidoOriana HernándezNo ratings yet
- Asfixia NeonatalDocument5 pagesAsfixia Neonatalnicaragua pediatrica100% (2)
- VIGASDocument40 pagesVIGASEdissonJavierNo ratings yet
- Bus - Especificaciones k410 6x2 Low Driver - tcm80-257452Document2 pagesBus - Especificaciones k410 6x2 Low Driver - tcm80-257452Juan Diego Chinchay Rojas100% (2)
- Sepsis Neonatal - Guía ClínicaDocument5 pagesSepsis Neonatal - Guía Clínicanicaragua pediatrica100% (5)
- Orden CerradoDocument44 pagesOrden CerradoOmar Alex Apaza MarcaNo ratings yet
- Síndrome de Reconstitución Inmune Por TuberculosisDocument2 pagesSíndrome de Reconstitución Inmune Por Tuberculosisnicaragua pediatricaNo ratings yet
- Un Gato Construye Una FamiliaDocument89 pagesUn Gato Construye Una FamiliaMika BrightNo ratings yet
- Dossier Beljanski PDFDocument24 pagesDossier Beljanski PDFMARC9249No ratings yet
- Revista NICARAGUA PEDIÁTRICA Septiembre-Diciembre ©2015Document42 pagesRevista NICARAGUA PEDIÁTRICA Septiembre-Diciembre ©2015nicaragua pediatrica100% (1)
- Revista NICARAGUA PEDIÁTRICA Mayo-Agosto ©2015Document46 pagesRevista NICARAGUA PEDIÁTRICA Mayo-Agosto ©2015nicaragua pediatrica100% (1)
- Prevalencia y Comportamiento Del Retinoblastoma en Niños Menores de 5 Años de EdadDocument5 pagesPrevalencia y Comportamiento Del Retinoblastoma en Niños Menores de 5 Años de Edadnicaragua pediatricaNo ratings yet
- Imagen Radiológica. Situs Inversus TotalisDocument1 pageImagen Radiológica. Situs Inversus Totalisnicaragua pediatricaNo ratings yet
- Manejo Quirúrgico de Las Complicaciones de HipospadiaDocument4 pagesManejo Quirúrgico de Las Complicaciones de Hipospadianicaragua pediatricaNo ratings yet
- Congresos 2015 ManaguaDocument1 pageCongresos 2015 Managuanicaragua pediatricaNo ratings yet
- BiografíaDocument1 pageBiografíanicaragua pediatricaNo ratings yet
- Displasia Tanatofórica Tipo IIDocument5 pagesDisplasia Tanatofórica Tipo IInicaragua pediatrica100% (1)
- Implicaciones de La Epigenética y La Regulación Del Estrés Sobre La Investigación y El DesarrolloDocument4 pagesImplicaciones de La Epigenética y La Regulación Del Estrés Sobre La Investigación y El Desarrollonicaragua pediatricaNo ratings yet
- Parámetros Utilizados en El Diagnóstico de Asfixia en El HMEADBDocument7 pagesParámetros Utilizados en El Diagnóstico de Asfixia en El HMEADBnicaragua pediatricaNo ratings yet
- Características Clínicas y Microbiológicas en Pacientes Hemato-Oncológicos Ingresados en El HIMJRDocument7 pagesCaracterísticas Clínicas y Microbiológicas en Pacientes Hemato-Oncológicos Ingresados en El HIMJRnicaragua pediatricaNo ratings yet
- Dr. Clarence Silva MartínezDocument2 pagesDr. Clarence Silva Martíneznicaragua pediatricaNo ratings yet
- EditorialDocument1 pageEditorialnicaragua pediatricaNo ratings yet
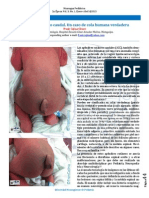
- Apéndice Cutáneo Caudal. Un Caso de Cola Humana VerdaderaDocument2 pagesApéndice Cutáneo Caudal. Un Caso de Cola Humana Verdaderanicaragua pediatricaNo ratings yet
- Editorial Mayo-Agosto 2015Document1 pageEditorial Mayo-Agosto 2015nicaragua pediatricaNo ratings yet
- Síndrome de Bridas AmnióticasDocument3 pagesSíndrome de Bridas Amnióticasnicaragua pediatricaNo ratings yet
- Características Epidemiológicas, Clínicas y Función Vesical en Niños (As) Con Vejiga NeurogénicaDocument5 pagesCaracterísticas Epidemiológicas, Clínicas y Función Vesical en Niños (As) Con Vejiga Neurogénicanicaragua pediatricaNo ratings yet
- Factores de Riesgo Asociados A Ictericia NeonatalDocument6 pagesFactores de Riesgo Asociados A Ictericia Neonatalnicaragua pediatrica67% (3)
- Efectividad Del Tratamiento Inmunosupresor Utilizado en El Manejo Del S. Nefrótico Corticorresistente y Su Relación Con La Variedad HistolDocument6 pagesEfectividad Del Tratamiento Inmunosupresor Utilizado en El Manejo Del S. Nefrótico Corticorresistente y Su Relación Con La Variedad Histolnicaragua pediatricaNo ratings yet
- Dr. Francisco Lacayo MaisonDocument2 pagesDr. Francisco Lacayo Maisonnicaragua pediatrica100% (1)
- Epigenética en PediatríaDocument5 pagesEpigenética en Pediatríanicaragua pediatrica100% (1)
- Comportamiento Clínico Quirúrgico de Los Pacientes Con Enfermedad de Hirschsprung, Que Fueron Intervenidos Mediante Técnica Mínimamente InvasivaDocument6 pagesComportamiento Clínico Quirúrgico de Los Pacientes Con Enfermedad de Hirschsprung, Que Fueron Intervenidos Mediante Técnica Mínimamente Invasivanicaragua pediatricaNo ratings yet
- Guía de Intervenciones Basada en Evidencias Que Reducen Morbilidad y Mortalidad Perinatal y NeonatalDocument144 pagesGuía de Intervenciones Basada en Evidencias Que Reducen Morbilidad y Mortalidad Perinatal y NeonatalElizabeth HenryNo ratings yet
- Comportamiento Epidemiológico de Los DC Más Frecuentes Del SNC en El HECAM 2004-2013Document6 pagesComportamiento Epidemiológico de Los DC Más Frecuentes Del SNC en El HECAM 2004-2013nicaragua pediatricaNo ratings yet
- ESTERILIZACIÓN POR MÉTODOS FÍSICOS Brat AcuacheDocument6 pagesESTERILIZACIÓN POR MÉTODOS FÍSICOS Brat AcuacheBrat AcuacheNo ratings yet
- Triptico Ergonomía Laboral 06.02Document2 pagesTriptico Ergonomía Laboral 06.02joseph elsen campos canchariNo ratings yet
- Dolphin 9900Document2 pagesDolphin 9900webcristiana100% (1)
- Fracturas Costales y FX de CaderaDocument56 pagesFracturas Costales y FX de CaderaXto Nuestra PazNo ratings yet
- Unidad Educativa Franciscana San AntonioDocument23 pagesUnidad Educativa Franciscana San Antoniomajo arellanoNo ratings yet
- Abraza Tu Poder InformeDocument12 pagesAbraza Tu Poder InformeGregorio InesNo ratings yet
- Hvac Memoria Descriptiva Costa AzulDocument19 pagesHvac Memoria Descriptiva Costa AzulJose MariaNo ratings yet
- Examen InicialDocument4 pagesExamen Inicialjose barreiroNo ratings yet
- Trabajo Monográfico-Vat Leaching. 1Document27 pagesTrabajo Monográfico-Vat Leaching. 1Rodrigo GuerraNo ratings yet
- Ou 2Document177 pagesOu 2Willy Ciprian PuenteNo ratings yet
- Faustina Rodríguez GutiérrezDocument85 pagesFaustina Rodríguez GutiérrezEric VázquezNo ratings yet
- IGLR - EspanholDocument40 pagesIGLR - EspanholAna Lúcia GomesNo ratings yet
- Solicitud RequerimientosDocument3 pagesSolicitud RequerimientosDino AndrésNo ratings yet
- Ingreso A ObrasDocument7 pagesIngreso A ObrasEugenio Alberto Neira VeraNo ratings yet
- Tamayo V 2018Document6 pagesTamayo V 2018Nadya FaizatuzNo ratings yet
- Anexo 1. Instrumento Autoevaluación PAPE (v1)Document64 pagesAnexo 1. Instrumento Autoevaluación PAPE (v1)Norelis Herrera CassianiNo ratings yet
- Cirque du Soleil: Arte y amor en mundos lejanosDocument6 pagesCirque du Soleil: Arte y amor en mundos lejanosNICOLLE QUEENNo ratings yet
- Ejemplo 2Document44 pagesEjemplo 2Yadira ChavezNo ratings yet
- Protocolo de Bioseguridad Itaa 2021 - V3Document32 pagesProtocolo de Bioseguridad Itaa 2021 - V3Yiseth MuñozNo ratings yet
- Inspección y pruebas de recipientes a presión soldadosDocument1 pageInspección y pruebas de recipientes a presión soldadosmguerra_635875No ratings yet
- Iiee-Metrados - (20.12.2021) Rev 01Document4 pagesIiee-Metrados - (20.12.2021) Rev 01Christiano RodriguezNo ratings yet
- Gestión Responsable de La Innovación (Mapa Conceptual) - JLDocument1 pageGestión Responsable de La Innovación (Mapa Conceptual) - JLJorge Eduardo Loayza Pérez100% (2)
- Mitos y Supersticiones PDFDocument312 pagesMitos y Supersticiones PDFMateo Palacio100% (1)