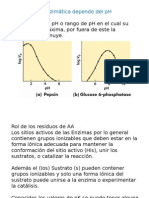
You might also like
- EnzimasDocument62 pagesEnzimasyurd leidy100% (4)
- EnzimasDocument10 pagesEnzimasvickyNo ratings yet
- Inhibicion EnzimaticaDocument14 pagesInhibicion EnzimaticaFiorella Rosas RuizNo ratings yet
- Enzimas ReguladorasDocument4 pagesEnzimas ReguladorasLucas Tapia Parra0% (1)
- Inhibidores EnzimáticosDocument9 pagesInhibidores EnzimáticosVariedades Y-JNo ratings yet
- Práctica 2. Determinación de Los Parámetros Cinéticos de La InvertasaDocument6 pagesPráctica 2. Determinación de Los Parámetros Cinéticos de La InvertasaMARTIN GARCIANo ratings yet
- CursoDocument37 pagesCursoMarco Antonio Salazar PalaciosNo ratings yet
- Qué Es La Cinética EnzimáticaDocument4 pagesQué Es La Cinética EnzimáticaJesús RGNo ratings yet
- Trabajo 3 de CineticaDocument9 pagesTrabajo 3 de Cineticachrismetal 188No ratings yet
- Ejercicios de Inhibición-Activación y Efectos de PHDocument3 pagesEjercicios de Inhibición-Activación y Efectos de PHaaron30102023No ratings yet
- Inhibidor EnzimáticoDocument18 pagesInhibidor EnzimáticoAdri CruzNo ratings yet
- Repaso de Cinética Enzimática e Inhibición Enzimática Fabián Rodríguez 2010Document4 pagesRepaso de Cinética Enzimática e Inhibición Enzimática Fabián Rodríguez 2010luis19401809No ratings yet
- Inhibición y Regulación EnzimáticaDocument5 pagesInhibición y Regulación EnzimáticaLourdes SoriaNo ratings yet
- Regulacion EnzimaticaDocument12 pagesRegulacion EnzimaticaKurtis JuarezNo ratings yet
- EnzimasDocument8 pagesEnzimasDavid JimenezNo ratings yet
- Qué Es Cinética EnzimáticaDocument21 pagesQué Es Cinética EnzimáticaStefa WupamNo ratings yet
- Propiedades Fundamentales de Una EnzimaDocument6 pagesPropiedades Fundamentales de Una Enzima•Estella Bemón•No ratings yet
- Tp02 de Bioquímica UNLPDocument11 pagesTp02 de Bioquímica UNLPAlyson GuilhermeNo ratings yet
- Tema 5 - EnzimasDocument14 pagesTema 5 - EnzimasSara DoradoNo ratings yet
- 6.3 BioquimicaDocument40 pages6.3 BioquimicaBrenda Soto100% (1)
- Definiciones QuimicaDocument5 pagesDefiniciones Quimicadenisse floresNo ratings yet
- Taller y Glosario de Enzimas y Factores Que Afectan Las EnzimasDocument12 pagesTaller y Glosario de Enzimas y Factores Que Afectan Las EnzimasAngie Villarreal100% (1)
- Aporte Tarea 3 BioquimicaDocument15 pagesAporte Tarea 3 BioquimicaSandra PeñaNo ratings yet
- Bioquimica Guia de Estudio 2Document21 pagesBioquimica Guia de Estudio 2daniel torresNo ratings yet
- Cinética EnzimaticaDocument23 pagesCinética EnzimaticaArturo MoraNo ratings yet
- Factores EnzimasDocument4 pagesFactores EnzimasMatiasHerreraNo ratings yet
- Cinética EnzimáticaDocument4 pagesCinética EnzimáticaDanii Castle RoblesNo ratings yet
- ME-Objetivos Enzimas RevDocument20 pagesME-Objetivos Enzimas Revapi-369665675% (4)
- Resúmen 3Document30 pagesResúmen 3Ramiro MartinNo ratings yet
- Efecto de La Concentracion de SustratoDocument9 pagesEfecto de La Concentracion de SustratoFatima SanchezNo ratings yet
- Bitacora 6Document4 pagesBitacora 6Morales Ramos PriscilaNo ratings yet
- Mecanismos de Union e Inhibición de EnzimasDocument7 pagesMecanismos de Union e Inhibición de EnzimasNALLELY GUADALUPE VITAL CARMONANo ratings yet
- Cinética EnzimáticaDocument7 pagesCinética EnzimáticaCelisdey CastilloNo ratings yet
- Resoluciön Ejercicios de Inhibición-Activación y Efectos de PHDocument2 pagesResoluciön Ejercicios de Inhibición-Activación y Efectos de PHaaron30102023No ratings yet
- Inhibidores PDFDocument7 pagesInhibidores PDFAnais FagundezNo ratings yet
- Info de Diapo Jorya AngelloDocument7 pagesInfo de Diapo Jorya AngelloJESUS ALBERTO GUERRERO DE LA ROSA ESTUDIANTE ACTIVONo ratings yet
- Enzimas IDocument6 pagesEnzimas IV PrNo ratings yet
- Documento Completo de Cinetica EnzimaticaDocument11 pagesDocumento Completo de Cinetica EnzimaticaMonica VegaNo ratings yet
- II Enzimología DiagnósticaDocument5 pagesII Enzimología DiagnósticaKaty LopezNo ratings yet
- Cuestionarios de BioquimicaDocument33 pagesCuestionarios de Bioquimicameme.gdjgtxNo ratings yet
- 5) Regulación de La Actividad Enzimática.: Figura 1. Enzimas ReguladorasDocument7 pages5) Regulación de La Actividad Enzimática.: Figura 1. Enzimas ReguladorasCortes Burgos Cielo SherlynNo ratings yet
- Enzimas 4Document48 pagesEnzimas 4MicuentoDehadasNo ratings yet
- Tarea No.9 Cinética EnzimáticaDocument8 pagesTarea No.9 Cinética EnzimáticaRodrigo ViguerasNo ratings yet
- Enzimas T6Document4 pagesEnzimas T6mzspqbzx7dNo ratings yet
- Inhibicion y SustratoDocument3 pagesInhibicion y SustratoYara HernandezNo ratings yet
- Lecture - 4 - QUÍMICA AMBIENTAL Y ELEMENTOS DE BIOQUÍMICA UNI IVDocument45 pagesLecture - 4 - QUÍMICA AMBIENTAL Y ELEMENTOS DE BIOQUÍMICA UNI IVfernando felipe reyes castilloNo ratings yet
- Bioquimica Biologia Celular Yt MolecularDocument89 pagesBioquimica Biologia Celular Yt MolecularGiulli DuránNo ratings yet
- Bioquimica 2023 TP 02 Enzimas - Preguntas Domiciliarias BIOQUIDocument8 pagesBioquimica 2023 TP 02 Enzimas - Preguntas Domiciliarias BIOQUIMati DPNo ratings yet
- Enzimas 2Document5 pagesEnzimas 2mpmNo ratings yet
- Inhibidores Enzimáticos de Uso ClínicosDocument5 pagesInhibidores Enzimáticos de Uso ClínicosAngélica RomanNo ratings yet
- Catálisis EnzimáticaDocument8 pagesCatálisis EnzimáticaMarta CastroNo ratings yet
- Práctica No 3 BioquimicaDocument9 pagesPráctica No 3 BioquimicaJesus E. Cabezudo CalderonNo ratings yet
- Arreglado para ImprimirDocument31 pagesArreglado para ImprimirMilton Cesar Rodrigo TantaleanNo ratings yet
- Inhibicion CompetitivaDocument9 pagesInhibicion Competitivakarla torrezNo ratings yet
- En ZimasDocument71 pagesEn ZimasLeidy ItzelNo ratings yet
- Clase 11 - Enzimas ReguladorasDocument81 pagesClase 11 - Enzimas ReguladorasJaime Saavedra Guevara100% (3)
- Ilovepdf - Merged (1) - MergedDocument190 pagesIlovepdf - Merged (1) - MergedMARIA FERNANDA SERVIN ROBLEDONo ratings yet
- Cuestionario ENZIMASDocument18 pagesCuestionario ENZIMASCristina Russell100% (1)
- Bbioquimica de Los Alimentos - Enzimas en Panaderia y ConfiteriaDocument33 pagesBbioquimica de Los Alimentos - Enzimas en Panaderia y ConfiteriaHenry Oré Gutiérrez83% (6)
- Enzimas Reguladoras o Enzimas AlostéricasDocument5 pagesEnzimas Reguladoras o Enzimas AlostéricasNitgma DcNo ratings yet
- Hematología MicrobiologiaDocument62 pagesHematología MicrobiologiaNegrita GuevaraNo ratings yet
- En La Investigación Cuantitativa WordDocument2 pagesEn La Investigación Cuantitativa WordNegrita GuevaraNo ratings yet
- LigandosDocument7 pagesLigandosNegrita GuevaraNo ratings yet
- HONGOSDocument3 pagesHONGOSNegrita GuevaraNo ratings yet
- AjedrezDocument8 pagesAjedrezNegrita GuevaraNo ratings yet
- Jugo GástricoDocument5 pagesJugo GástricoNegrita GuevaraNo ratings yet
- Prueba de CatalasaDocument3 pagesPrueba de CatalasaNegrita Guevara100% (1)
- NEFELÓMETRODocument2 pagesNEFELÓMETRONegrita Guevara0% (1)
- Ejercicios Enzimas PDFDocument3 pagesEjercicios Enzimas PDFDavid AntonioNo ratings yet
- GPT (Alt) : Determinación Cuantitativa de Alanina Aminotransferasa GPT (Alt)Document1 pageGPT (Alt) : Determinación Cuantitativa de Alanina Aminotransferasa GPT (Alt)FernandaNo ratings yet
- Citocromo p450Document17 pagesCitocromo p450Natalia BahamonNo ratings yet
- EnzimasDocument5 pagesEnzimasMaría Belén Hernández OrtizNo ratings yet
- Aspectos Generales y Propiedades de Las EnzimasDocument37 pagesAspectos Generales y Propiedades de Las Enzimasronald stevn alvarez osorioNo ratings yet
- EnzimasDocument13 pagesEnzimasMariana CastilloNo ratings yet
- 5) Regulación de La Actividad Enzimática.: Figura 1. Enzimas ReguladorasDocument7 pages5) Regulación de La Actividad Enzimática.: Figura 1. Enzimas ReguladorasCortes Burgos Cielo SherlynNo ratings yet
- Mapa Mental BiocaDocument1 pageMapa Mental BiocaAna ReyesNo ratings yet
- Modificación de La Actividad Enzimática de La TirosinasaDocument28 pagesModificación de La Actividad Enzimática de La TirosinasaDaniela VeraNo ratings yet
- Tabla Comparativa de Inhibición EnzimáticaDocument2 pagesTabla Comparativa de Inhibición EnzimáticaLizbeth RamirezNo ratings yet
- ENZIMIOLOGÍA - Obstetricia PDFDocument65 pagesENZIMIOLOGÍA - Obstetricia PDFDerecho IcaNo ratings yet
- ENZIMAS Capitulo 13 BISHOPDocument25 pagesENZIMAS Capitulo 13 BISHOPCRISTIAN EDUARDO MARTINEZ DAVILANo ratings yet
- Problemas de Bioquímica - Primer Curso Del Grado en Bioquímica - Curso 2010 11Document8 pagesProblemas de Bioquímica - Primer Curso Del Grado en Bioquímica - Curso 2010 11cristipisti88No ratings yet
- Metabolismo Del GlucogenoDocument23 pagesMetabolismo Del GlucogenoFeer RangelNo ratings yet
- Reaccion de TransaminacionDocument2 pagesReaccion de TransaminacionXkevin ZNo ratings yet
- Bioquimica Basica Base Molecular de Los Procesos Fisiologicos Cap 3Document20 pagesBioquimica Basica Base Molecular de Los Procesos Fisiologicos Cap 3Elda PasosNo ratings yet
- Enzimas. Enzimología ClínicaDocument39 pagesEnzimas. Enzimología ClínicaAlexNo ratings yet
- Rodriguez - MariaJose - Tarea 2.1Document4 pagesRodriguez - MariaJose - Tarea 2.1Majo RodriguezNo ratings yet
- Clase 6 Control de La Actividad EnzimáticaDocument36 pagesClase 6 Control de La Actividad EnzimáticaHarold JoseNo ratings yet
- Presentación EnzimasDocument33 pagesPresentación EnzimasAndrea HinestrozaNo ratings yet
- Original Trabajo de EnzimaDocument22 pagesOriginal Trabajo de Enzimadaysi raquel florez vertelNo ratings yet
- Enzimologia ClinicaDocument18 pagesEnzimologia ClinicaRoberto Castellanos100% (1)
- Tiosulfonil TransferasaDocument3 pagesTiosulfonil TransferasaJorge Ramirez RaherNo ratings yet
- Cardona - Los Enzimas. Introducción A La Enzimología.Document9 pagesCardona - Los Enzimas. Introducción A La Enzimología.Danher EscalanteNo ratings yet
- Semana 4 EnzimasDocument65 pagesSemana 4 EnzimasYassimNo ratings yet
- EXTRACCIÓN DE FOSFATASA ALCALINA, EFECTO DE PH, TEMPERATURA, VELOCIDAD DE REACCIÓN Y ACTIVIDAD ENZIMÁTICADocument6 pagesEXTRACCIÓN DE FOSFATASA ALCALINA, EFECTO DE PH, TEMPERATURA, VELOCIDAD DE REACCIÓN Y ACTIVIDAD ENZIMÁTICAa.agudelo2No ratings yet
- Unidad 3 Enzimas - Parte IDocument70 pagesUnidad 3 Enzimas - Parte IYony RMNo ratings yet
- AlosteríaDocument1 pageAlosteríaDaniela MNo ratings yet
- Ciclo de KrebsDocument2 pagesCiclo de Krebsshin e izanely The hedgehogNo ratings yet