You might also like
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Cell Organelles and Their FunctionsDocument28 pagesCell Organelles and Their FunctionsKayeNo ratings yet
- Death Watch Beetle: in CultureDocument2 pagesDeath Watch Beetle: in Culturewinofvin9No ratings yet
- 670300-3 Investigation of Starling Populations in BCDocument57 pages670300-3 Investigation of Starling Populations in BCGbah ChanNo ratings yet
- Normal Radiographical AnatomyDocument30 pagesNormal Radiographical AnatomyALI abd-alamamNo ratings yet
- Propose A SolutionDocument21 pagesPropose A SolutionPhước NguyễnNo ratings yet
- Cell DivisionDocument72 pagesCell Divisionnor ainiNo ratings yet
- Muscles of MasticationDocument18 pagesMuscles of MasticationgauravNo ratings yet
- Should We Eat BugsDocument2 pagesShould We Eat BugsbradylynNo ratings yet
- Science 10 Bio WorksheetsDocument15 pagesScience 10 Bio WorksheetsMemmy KidnaNo ratings yet
- Overview of Hominin EvolutionDocument5 pagesOverview of Hominin EvolutionJaviera GibertoNo ratings yet
- B.SC 1st Yr Syl. 101 & 102-1Document3 pagesB.SC 1st Yr Syl. 101 & 102-1Anwita JhaNo ratings yet
- Histology of Digestive SystemDocument100 pagesHistology of Digestive SystemFadhila Putri Palupi100% (3)
- Histo 1st ShiftDocument25 pagesHisto 1st ShiftIvy CruzNo ratings yet
- Powerpoint Lectures For: Biology, Seventh EditionDocument124 pagesPowerpoint Lectures For: Biology, Seventh Editionxo_simpledream100% (1)
- Vertebrates and Invertebrates. Science 5esquemaDocument1 pageVertebrates and Invertebrates. Science 5esquemaBLANCA MASERONo ratings yet
- Plant StructureDocument6 pagesPlant StructureYoga Trista MajidNo ratings yet
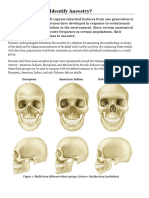
- Activity: Can You Identify Ancestry?: European American Indian Sub-Saharan AfricanDocument5 pagesActivity: Can You Identify Ancestry?: European American Indian Sub-Saharan AfricanpoovenmoonoosamyNo ratings yet
- Stylelife - The Red Queen Study GuideDocument7 pagesStylelife - The Red Queen Study GuideNewStarGodNo ratings yet
- Plant ReproductionDocument11 pagesPlant ReproductionRagavNarayanan0% (1)
- Patterning of DentitionDocument4 pagesPatterning of Dentitionimi4No ratings yet
- Unit 1 Living Things Natural Science 3ºlast PDFDocument34 pagesUnit 1 Living Things Natural Science 3ºlast PDFJosefa Jerez0% (1)
- Excretory Products and Their EliminationDocument9 pagesExcretory Products and Their EliminationBiju MylachalNo ratings yet
- Colegio de San Juan de LetranDocument71 pagesColegio de San Juan de Letranirish xNo ratings yet
- Introduction To The HelminthsDocument7 pagesIntroduction To The HelminthsmuftizamanNo ratings yet
- Npre2007241 2 PDFDocument134 pagesNpre2007241 2 PDFranjeet kumarNo ratings yet
- 21 Sargetia - Acta Musei Devensis - Series Scientia Naturae XXI 2008 PDFDocument185 pages21 Sargetia - Acta Musei Devensis - Series Scientia Naturae XXI 2008 PDFMaria TurcuNo ratings yet
- Vocabulary: Animal QuizDocument1 pageVocabulary: Animal QuizMariciell Larissa Gonzales TiconaNo ratings yet
- Critical Thinking On Certain Questions Concerning Comparative Anatomy of The VertebratesDocument5 pagesCritical Thinking On Certain Questions Concerning Comparative Anatomy of The VertebratesSeth Andrew SalihNo ratings yet
- Captive Care of North American Water TurtlesDocument160 pagesCaptive Care of North American Water TurtlesAnthony Lopez SahagunNo ratings yet
- Exploring Animal Life Cycles: Ms. Mckeel & Ms. Messina 2nd GradeDocument15 pagesExploring Animal Life Cycles: Ms. Mckeel & Ms. Messina 2nd Gradeapi-302056293No ratings yet