You might also like
- Komunikasi Antar Sel Menggunakan Molekul Signal EkstraselulerDocument14 pagesKomunikasi Antar Sel Menggunakan Molekul Signal EkstraselulerAfif Al Fatih100% (1)
- KOMUNIKASI ANTAR SELDocument11 pagesKOMUNIKASI ANTAR SELice novriNo ratings yet
- SELMENDocument14 pagesSELMENErmawati Erma100% (2)
- Komunikasi SelDocument15 pagesKomunikasi SelussyNo ratings yet
- Komunikasi SelDocument2 pagesKomunikasi SelCitra Nur MutiarahmiNo ratings yet
- Cara Sel Berkomunikasi Untuk Mempertahankan Homeostasis Dan TumbuhDocument1 pageCara Sel Berkomunikasi Untuk Mempertahankan Homeostasis Dan TumbuhRamadhani Syafitri100% (1)
- Cell SignallingDocument50 pagesCell SignallingDuwi EfasariNo ratings yet
- Mekanisme Transpor Membran PlasmaDocument3 pagesMekanisme Transpor Membran PlasmareafyNo ratings yet
- Interaksi Dan Komunikasi Antar SelDocument25 pagesInteraksi Dan Komunikasi Antar Selaverizkaqueen100% (7)
- HCG FUNGSI BIOLOGIS DAN APLIKASI KLINISDocument12 pagesHCG FUNGSI BIOLOGIS DAN APLIKASI KLINISelNo ratings yet
- Cell Junction TugasDocument6 pagesCell Junction TugasStefany LukeNo ratings yet
- Struktur Fungsi EndomembranDocument22 pagesStruktur Fungsi EndomembranEstu Ria Dwi YulianingsihNo ratings yet
- Ultrastruktur Sel - PPDS 2019Document33 pagesUltrastruktur Sel - PPDS 2019DudungNo ratings yet
- Klasifikasi Hormon LokalDocument3 pagesKlasifikasi Hormon LokalPutriArifiana100% (1)
- Sintesis Glikoprotein Dan ProteoglikanDocument6 pagesSintesis Glikoprotein Dan ProteoglikanrullNo ratings yet
- KOMSELSIGDocument13 pagesKOMSELSIGNurul Muhlisa MusNo ratings yet
- Kerja Hormon Dan Tranduksi SinyalDocument23 pagesKerja Hormon Dan Tranduksi SinyalQd Vella QnozesDanteNo ratings yet
- Komunikasi SelDocument22 pagesKomunikasi SelRaudhah RamadiyantikaNo ratings yet
- Serius Proses Homeostasis Pada Tubuh BerkeringatDocument9 pagesSerius Proses Homeostasis Pada Tubuh BerkeringatYussi Septiana100% (2)
- Antigen AntibodiDocument32 pagesAntigen AntibodiAriep SeduniaNo ratings yet
- BIOKIMIA MEMBRANDocument41 pagesBIOKIMIA MEMBRANZhafran DarwisNo ratings yet
- Komunikasi SelDocument7 pagesKomunikasi SelAndrio PalayukanNo ratings yet
- TutorDocument6 pagesTutorMort KowalskiNo ratings yet
- PATOFISIOLOGI DM TIPE 1 DAN 2Document2 pagesPATOFISIOLOGI DM TIPE 1 DAN 2Fiddiyah Galuh50% (2)
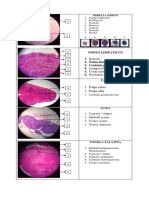
- Medulla ossium dan organ limfatikDocument1 pageMedulla ossium dan organ limfatikMerlintaNo ratings yet
- KARIOTIPE DAN ANEUPLOIDYDocument14 pagesKARIOTIPE DAN ANEUPLOIDYRaudlatul_19jannahNo ratings yet
- P53Document11 pagesP53Ika SukmaNo ratings yet
- HDN-PENDAHULUANDocument21 pagesHDN-PENDAHULUANRizky PatimahNo ratings yet
- Apoptosis Dan NekrosisDocument7 pagesApoptosis Dan NekrosisNaditya Fitriani HasanahNo ratings yet
- Mekanisme Kerja Hormon SteroidDocument6 pagesMekanisme Kerja Hormon SteroidRoy Befi100% (6)
- KOMUNIKASI SELDocument13 pagesKOMUNIKASI SELsuci_cewexmanies100% (2)
- Makalah Siklus SelDocument9 pagesMakalah Siklus SelAinun Zaskia SyamsuNo ratings yet
- ALOSTERIKDocument2 pagesALOSTERIKhendra_padang100% (2)
- Proses Sintesis ProteinDocument31 pagesProses Sintesis Proteinmaudy octhalia d4 kepNo ratings yet
- Etiologi JejasDocument15 pagesEtiologi JejasYulia Dewi PratiwiNo ratings yet
- Komunikasi SelDocument44 pagesKomunikasi SelKiaNo ratings yet
- Sistem Pencernaan: Usus Halus dan Usus BesarDocument22 pagesSistem Pencernaan: Usus Halus dan Usus BesarJim KhulafaNo ratings yet
- Analisis KromosomDocument4 pagesAnalisis KromosomdeyaNo ratings yet
- Ion Kalsium Pada Metabolisme Matriks EkstrselulerDocument37 pagesIon Kalsium Pada Metabolisme Matriks EkstrselulerKris RatuNo ratings yet
- Hukum Starling Pada Laju Filtrasi GlomerulusDocument1 pageHukum Starling Pada Laju Filtrasi GlomerulusZyad Kemal0% (1)
- Antigen Dan Struktur AntigenDocument17 pagesAntigen Dan Struktur AntigenMADU BASUKINo ratings yet
- KONSEP RESEPTOR DAN SEJARAHNYADocument38 pagesKONSEP RESEPTOR DAN SEJARAHNYARizmá RizemNo ratings yet
- Makalah HomeostatisDocument23 pagesMakalah Homeostatismulkihakam21No ratings yet
- Laporan Praktikum (Gabungan)Document90 pagesLaporan Praktikum (Gabungan)Novia KaisariantiNo ratings yet
- Makalah GlukoneogenesisDocument31 pagesMakalah GlukoneogenesisIkhwan Latif0% (1)
- Anatomi Fisiologi Sistem EndokrinDocument14 pagesAnatomi Fisiologi Sistem EndokrinUMan NugrrozNo ratings yet
- Sistem ImunDocument16 pagesSistem ImunAbdillah AriefNo ratings yet
- KARBOHIDRAT DAN NUKLEOTIDADocument75 pagesKARBOHIDRAT DAN NUKLEOTIDAAndi Fitria RizkitaNo ratings yet
- IMUNITAS PARUDocument13 pagesIMUNITAS PARUMuhammad AriefNo ratings yet
- Makalah SignalingDocument16 pagesMakalah SignalingCut IndriputriNo ratings yet
- Rosalina Yolanda - 020.06.0073 - B (Essay Konsep Homeostasis Tubuh Pada Perubahan Kondisi Lingkungan Di Daerah Wisata)Document5 pagesRosalina Yolanda - 020.06.0073 - B (Essay Konsep Homeostasis Tubuh Pada Perubahan Kondisi Lingkungan Di Daerah Wisata)Rosalina YolandaNo ratings yet
- Biokimia-Ayu Puspitasari-Metabolisme Purin & Pirimidin 2014 PDFDocument18 pagesBiokimia-Ayu Puspitasari-Metabolisme Purin & Pirimidin 2014 PDFAnonymous Uh9XtlGN100% (5)
- Mekanisme ToksikologiDocument7 pagesMekanisme ToksikologiPenca AkunaNo ratings yet
- Histologi Sistem EndokrinDocument50 pagesHistologi Sistem Endokrindebbyelvira50% (2)
- BIOELEKTRIK JANTUNGDocument44 pagesBIOELEKTRIK JANTUNGAndreas OctavianoNo ratings yet
- MAKALAH Sistem Transport MembranDocument15 pagesMAKALAH Sistem Transport MembranTikaUlam100% (1)
- Optimized Essay Biology QuestionsDocument5 pagesOptimized Essay Biology QuestionsFadli MermangNo ratings yet
- Biokima CardiovaskularDocument28 pagesBiokima CardiovaskularAanNo ratings yet
- Transduksi SinyalDocument45 pagesTransduksi SinyalChandraNo ratings yet
- Makalah Komunikasi Antar SelDocument14 pagesMakalah Komunikasi Antar SelUnny Ru100% (2)
- Surat KuasaDocument1 pageSurat KuasaDoni HardiansyahNo ratings yet
- Skin CareDocument11 pagesSkin CareNunik Utari NurwulandariNo ratings yet
- SOP IZIN Surat Izin ApotekDocument2 pagesSOP IZIN Surat Izin ApotekNunik Utari NurwulandariNo ratings yet
- Format Surat Pernyataan Profil ApotekDocument1 pageFormat Surat Pernyataan Profil ApotekdoitaNo ratings yet
- KulitDocument18 pagesKulitNunik Utari NurwulandariNo ratings yet
- Pemasangan OksigenDocument7 pagesPemasangan OksigenNunik Utari NurwulandariNo ratings yet
- 012 Spo Telaah ResepDocument2 pages012 Spo Telaah ResepNunik Utari NurwulandariNo ratings yet
- Surat Pernyataan PC Surabaya OkDocument2 pagesSurat Pernyataan PC Surabaya OkNunik Utari NurwulandariNo ratings yet
- 018 Spo Penarikan ObatDocument3 pages018 Spo Penarikan ObatNunik Utari NurwulandariNo ratings yet
- KUISIONER Sebaarr TerakhirDocument5 pagesKUISIONER Sebaarr TerakhirNunik Utari NurwulandariNo ratings yet
- Rencana Pelaksanaan Pembelajaran CoverDocument1 pageRencana Pelaksanaan Pembelajaran CoverNunik Utari NurwulandariNo ratings yet
- Kimed Analgetik Non SteroidDocument3 pagesKimed Analgetik Non SteroidNunik Utari NurwulandariNo ratings yet
- Pengaruh Kombinasi Magnesium Stearat Dan Talkum Sebagai Lubrikan Terhadap Profil Disolusi Tablet IbuprofenDocument12 pagesPengaruh Kombinasi Magnesium Stearat Dan Talkum Sebagai Lubrikan Terhadap Profil Disolusi Tablet IbuprofenHidmatNo ratings yet
- Jadwal PembelajaranDocument1 pageJadwal PembelajaranNunik Utari NurwulandariNo ratings yet
- Daftar HadirDocument1 pageDaftar HadirNunik Utari NurwulandariNo ratings yet
- Hanya Cover OkeDocument2 pagesHanya Cover OkeNunik Utari NurwulandariNo ratings yet
- II.A. DDS OralDocument48 pagesII.A. DDS OralNunik Utari NurwulandariNo ratings yet
- RambutDocument17 pagesRambutNunik Utari NurwulandariNo ratings yet
- Pendahuluan KrimDocument12 pagesPendahuluan Krimrizki lisya nugrahaNo ratings yet
- Uu Nomor 33 Tahun 2014 Tentang Jaminan Produk HalalDocument27 pagesUu Nomor 33 Tahun 2014 Tentang Jaminan Produk HalalM Anton AthoillahNo ratings yet
- LEMAK DAN HIPERLIPIDEMIADocument13 pagesLEMAK DAN HIPERLIPIDEMIANunik Utari NurwulandariNo ratings yet
- KepadaDocument1 pageKepadaNunik Utari NurwulandariNo ratings yet
- OPTIMASI PENGHANTARAN MUKOSALDocument14 pagesOPTIMASI PENGHANTARAN MUKOSALNunik Utari NurwulandariNo ratings yet
- Pengaruh Kombinasi Magnesium Stearat Dan Talkum Sebagai Lubrikan Terhadap Profil Disolusi Tablet IbuprofenDocument12 pagesPengaruh Kombinasi Magnesium Stearat Dan Talkum Sebagai Lubrikan Terhadap Profil Disolusi Tablet IbuprofenHidmatNo ratings yet
- Bab VDocument4 pagesBab VNunik Utari NurwulandariNo ratings yet
- Kriteria KKMDocument9 pagesKriteria KKMNunik Utari NurwulandariNo ratings yet
- Lampiran 1 Strategi PelaksanaanDocument12 pagesLampiran 1 Strategi PelaksanaanNunik Utari NurwulandariNo ratings yet
- Daftar PustakaDocument1 pageDaftar PustakaNunik Utari NurwulandariNo ratings yet
- Kata PengantarDocument3 pagesKata PengantarNunik Utari NurwulandariNo ratings yet
- Daftar IsiDocument3 pagesDaftar IsiNunik Utari NurwulandariNo ratings yet