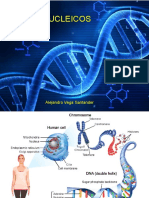
You might also like
- Historia y desarrollo de la citogenética convencionalDocument38 pagesHistoria y desarrollo de la citogenética convencionalClaudia IpuchaNo ratings yet
- Clase de Citogenética HumanaDocument62 pagesClase de Citogenética HumanaPaola FloresNo ratings yet
- Biologia Mapas GeneticosDocument24 pagesBiologia Mapas GeneticosPerla Liliana100% (1)
- CitogeneticaDocument96 pagesCitogeneticaOvielma100% (1)
- CitogeneticaDocument192 pagesCitogeneticaholmangaitanNo ratings yet
- Factores Sigma en Escherichia ColiDocument4 pagesFactores Sigma en Escherichia ColiEncb IpnNo ratings yet
- 5ta Clase Genetica Ligamiento y Recombinación de GenesDocument9 pages5ta Clase Genetica Ligamiento y Recombinación de GenesRena HenriquezNo ratings yet
- ClonaciónDocument6 pagesClonaciónLucho AccoNo ratings yet
- HISTOLOGIADocument41 pagesHISTOLOGIAPepe DelgadoNo ratings yet
- 4-Genetica MendelianaDocument22 pages4-Genetica MendelianaCaro PulidoNo ratings yet
- Ejerciicos de GeneticaDocument8 pagesEjerciicos de GeneticaDAVID IVAN RUIZ SILVA100% (1)
- Ejercicios GenéticaDocument2 pagesEjercicios GenéticaJaime de la MazaNo ratings yet
- 1-07 Acidos NucléicosDocument55 pages1-07 Acidos NucléicosDan EmpleoNo ratings yet
- Evolucion Sistematica y Taxonomia MicrobianaDocument33 pagesEvolucion Sistematica y Taxonomia Microbianaeliza humpiriNo ratings yet
- Leyes de Mendel y MásDocument34 pagesLeyes de Mendel y MásLiana Henderson MarreroNo ratings yet
- Imagenes 2do Solemne PracticoDocument8 pagesImagenes 2do Solemne PracticoCristobal CernaNo ratings yet
- Transcripción Del ADNDocument45 pagesTranscripción Del ADNClauMcClauNo ratings yet
- Problemas de Genética - CICLO II SECCIÓN G Ingeniería AmbientalDocument6 pagesProblemas de Genética - CICLO II SECCIÓN G Ingeniería AmbientalAngie Evita Reyes HualcasNo ratings yet
- B34 Parte1Document195 pagesB34 Parte1celestecalipsoNo ratings yet
- Anomalías genéticas de las proteínas estructuralesDocument41 pagesAnomalías genéticas de las proteínas estructuralesKatherine Niño AcevedoNo ratings yet
- Practica 8 MitocondriaDocument5 pagesPractica 8 MitocondriaDavid Camilo Alvarez RealNo ratings yet
- Reacciones FebrilesDocument2 pagesReacciones Febrilesgallito2406No ratings yet
- Urquiza Carezzano Morfologia Animal Tomo I GratisDocument163 pagesUrquiza Carezzano Morfologia Animal Tomo I GratisSergio Urquiza100% (1)
- Manual de Citogenetica Humana N. Wiener 2010Document41 pagesManual de Citogenetica Humana N. Wiener 2010jaquelinefp100% (1)
- Genética de Poblaciones: Ejercicios Alelos Ligados al SexoDocument2 pagesGenética de Poblaciones: Ejercicios Alelos Ligados al SexoBrianm FlorezNo ratings yet
- Biologia y Jurassic ParkDocument3 pagesBiologia y Jurassic ParkCarlos Chicunguya100% (1)
- Embriologia Veterinaria Jose Monterde yDocument183 pagesEmbriologia Veterinaria Jose Monterde yMariel Garcia AyalaNo ratings yet
- Análisis citogenético de plantasDocument75 pagesAnálisis citogenético de plantasJoselyn KarinaNo ratings yet
- Variaciones genéticas en procariotasDocument15 pagesVariaciones genéticas en procariotasRalphy Guanipa BarriosNo ratings yet
- Atlas Histológico Del Aparato Reproductor en MamíferosDocument31 pagesAtlas Histológico Del Aparato Reproductor en Mamíferosalvarocss89No ratings yet
- Problemas Resueltos de Genética 2Document21 pagesProblemas Resueltos de Genética 2Irene Ortega100% (1)
- Fundamentos de genéticaDocument45 pagesFundamentos de genéticaKevin GutierrezNo ratings yet
- Sistemas y Aparatos Del Cuerpo HumanoDocument37 pagesSistemas y Aparatos Del Cuerpo HumanoGisela Pino PizáNo ratings yet
- Semana 8. HongosDocument35 pagesSemana 8. HongosangieNo ratings yet
- Organizacion Del Material GeneticoDocument39 pagesOrganizacion Del Material GeneticoBelen Lorena Alvarez VargasNo ratings yet
- MARCADORES MOLECULARES Breve Revisión de Los Marcadores MolecularesDocument26 pagesMARCADORES MOLECULARES Breve Revisión de Los Marcadores MolecularesDavid Glez ZNo ratings yet
- Informe CultivosDocument10 pagesInforme CultivosGilson ValenciaNo ratings yet
- Glosario Biología MolecularDocument7 pagesGlosario Biología MolecularValeria100% (1)
- Atlas de HematologiaDocument22 pagesAtlas de HematologiaNazia EstelaNo ratings yet
- Antoine LavoisierDocument4 pagesAntoine LavoisierIsabel Cerecedo CurielNo ratings yet
- Mini Atlas - Série Leucocitárias - 1 EdiciónDocument137 pagesMini Atlas - Série Leucocitárias - 1 EdiciónJerzy AmbrizNo ratings yet
- Genetica BacterianaDocument19 pagesGenetica BacterianaJoel Ramirez PadillaNo ratings yet
- Ejercicios de AnomalíasDocument28 pagesEjercicios de AnomalíasEdgar Huamanchumo100% (1)
- Informe de SangreDocument7 pagesInforme de SangreSheila M DiazNo ratings yet
- Herencia y Genetica PDFDocument67 pagesHerencia y Genetica PDFCarla PegueroNo ratings yet
- Genética de PoblacionesDocument54 pagesGenética de PoblacionesSthefany GonzalesNo ratings yet
- Síntesis de Proteínas ExposicionDocument19 pagesSíntesis de Proteínas ExposicionCarl StanleyNo ratings yet
- CITOGENÉTICA Kane PDFDocument148 pagesCITOGENÉTICA Kane PDFKeiko Katerine Kanematsu Grados0% (1)
- MetabolismoDocument27 pagesMetabolismoLibros Andrea100% (2)
- Manual de ProblemasDocument160 pagesManual de ProblemasRakel Dreaming Inblue0% (1)
- Biología MolecularDocument46 pagesBiología MolecularGonzo TorpedoNo ratings yet
- Infografía ReplicaciónDocument3 pagesInfografía ReplicaciónBraulio HetfieldNo ratings yet
- Metabolismo Glucosa 1Document56 pagesMetabolismo Glucosa 1Moonie QuiñonesNo ratings yet
- Cariotipo Humano MonografíaDocument19 pagesCariotipo Humano MonografíaYvon Becerra CuevaNo ratings yet
- Citogenética Humana Oncohematologia PDFDocument8 pagesCitogenética Humana Oncohematologia PDFjhonandyramosNo ratings yet
- Cariotipo y Bandeo CDocument3 pagesCariotipo y Bandeo CMark AlexanderNo ratings yet
- Metodo de Bandeo CromosomicoDocument19 pagesMetodo de Bandeo Cromosomicoeldaquispesanchez100% (1)
- Bandeo CromosómicoDocument5 pagesBandeo CromosómicoErick Leonardo Valle Mendoza33% (3)
- Practica 3 Elaboración Del Cariograma DMSHDocument48 pagesPractica 3 Elaboración Del Cariograma DMSHpatriciaNo ratings yet
- Lengua - de .Señas - ArgentinaDocument27 pagesLengua - de .Señas - ArgentinaMaria Taborda50% (2)
- Piensa en Haskell - AlonsoJimenez&HidalgoDoblado - 2012Document428 pagesPiensa en Haskell - AlonsoJimenez&HidalgoDoblado - 2012EAPNo ratings yet
- La Deriva Errónea de La Inteligencia ArtificialDocument135 pagesLa Deriva Errónea de La Inteligencia ArtificialA6atc7No ratings yet
- WBS Curso 2011Document50 pagesWBS Curso 2011EAPNo ratings yet
- Casasus Pablo Neruda en Morelos 1941 1966Document97 pagesCasasus Pablo Neruda en Morelos 1941 1966Lindsay Vargas100% (1)
- La Nueva UtopiaDocument14 pagesLa Nueva UtopiaEAPNo ratings yet
- El imperialismo cultural y los pueblos de América LatinaDocument275 pagesEl imperialismo cultural y los pueblos de América LatinaManuel GonzálezNo ratings yet
- Smith Magenis Lab 2012Document26 pagesSmith Magenis Lab 2012EAPNo ratings yet
- Aristoteles ObraBiologicaDocument232 pagesAristoteles ObraBiologicaEAPNo ratings yet
- Eva PeronDocument81 pagesEva PeronEAP50% (2)
- David Bravo - Copia Este Libro PDFDocument144 pagesDavid Bravo - Copia Este Libro PDFDani KardoneNo ratings yet
- La Rebelión de Los Luditas (1982-2012)Document24 pagesLa Rebelión de Los Luditas (1982-2012)Héctor PeñaNo ratings yet
- El Oenegismo Enfermedad Infantil Del Derechismo Por Alvaro Garcia Linera-1Document168 pagesEl Oenegismo Enfermedad Infantil Del Derechismo Por Alvaro Garcia Linera-1Blanche TorricoNo ratings yet
- Embellecimiento Fraude y FalsificacionDocument121 pagesEmbellecimiento Fraude y FalsificacionEAP100% (1)
- Las convicciones de Erich FrommDocument253 pagesLas convicciones de Erich FrommJuan Pablo GaydouNo ratings yet
- Cómo Escribir y Publicar Trabajos CientíficosDocument270 pagesCómo Escribir y Publicar Trabajos CientíficosEstudiantes por Derecho88% (8)
- Las Moradas Filosofales - FulcanelliDocument221 pagesLas Moradas Filosofales - FulcanelliEl Jinete Pálido98% (46)
- Manejo SMSDocument10 pagesManejo SMSEAPNo ratings yet
- Guia de Enf RarasDocument108 pagesGuia de Enf RarasLorena SilvaNo ratings yet
- Primera Jornada de Difusión Del Síndrome de Smith-MagenisDocument1 pagePrimera Jornada de Difusión Del Síndrome de Smith-MagenisEAPNo ratings yet
- Internet Hackers Software LibreDocument242 pagesInternet Hackers Software LibreVotaguz94% (36)
- Python para TodosDocument160 pagesPython para TodosJunior Sumosa100% (2)
- Planeta Web2Document162 pagesPlaneta Web2anon-89343894% (17)
- Ni Más Ni Menos. La Discapacidad Explicada A Los Niños y NiñasDocument24 pagesNi Más Ni Menos. La Discapacidad Explicada A Los Niños y Niñasorientacionccv100% (2)
- El Síndrome de Smith-MagenisDocument8 pagesEl Síndrome de Smith-MagenisEAPNo ratings yet
- El Volcán LatinoamericanoDocument228 pagesEl Volcán LatinoamericanoFernando M. Galeana100% (2)
- Cariotipo, Trabajo PracticoDocument11 pagesCariotipo, Trabajo PracticoWendy DavilaNo ratings yet
- Síndrome de DiGeorgeDocument17 pagesSíndrome de DiGeorgeSusanaNo ratings yet
- La Biologia Molecular en La Oncohematologia 2020-2Document42 pagesLa Biologia Molecular en La Oncohematologia 2020-2LUZ MAGDALENA AVILA MECANo ratings yet
- Practica de CariotipoDocument7 pagesPractica de CariotipoEzequiel LopezNo ratings yet
- 750901474.SG 6 Cromosomopatías IDocument22 pages750901474.SG 6 Cromosomopatías ICATALINA ROMERO LERETENo ratings yet
- MosaicismoDocument3 pagesMosaicismoMainDroidNo ratings yet
- Biología celular: Cromosomas y ciclo celularDocument17 pagesBiología celular: Cromosomas y ciclo celularGian LuccaNo ratings yet
- Cromosomopatias. Alteraciones Numericas.Document57 pagesCromosomopatias. Alteraciones Numericas.Maria Jose Garcia BermejoNo ratings yet
- Preguntas para Analizar - Unidad 3 - Genética - Tarea 4.Document3 pagesPreguntas para Analizar - Unidad 3 - Genética - Tarea 4.Sofia MejiaNo ratings yet
- Técnicas CitogenéticasDocument12 pagesTécnicas CitogenéticasMaría Ángel Suárez HenriquezNo ratings yet
- Normas laboratorios 2017Document79 pagesNormas laboratorios 2017Pilar Rubi Trinidad RodriguezNo ratings yet
- AneuploidíaDocument7 pagesAneuploidíaFLOR DE MARIA ROSARIO HUARCAYA LIMACHINo ratings yet
- Anomalias Estructurales Del CromosomaDocument12 pagesAnomalias Estructurales Del CromosomaleocomputerNo ratings yet
- El Estudio de Los Cromosomas HumanosDocument11 pagesEl Estudio de Los Cromosomas HumanosBárbara LeiteNo ratings yet
- Aberraciones CromosómicasDocument34 pagesAberraciones CromosómicasDianiiz GutierrezNo ratings yet
- Genetica 3 ApunteDocument39 pagesGenetica 3 ApunteDaniela MuñozNo ratings yet
- EMBRIOLOGÍA - Guía de Práctica 2017Document81 pagesEMBRIOLOGÍA - Guía de Práctica 2017Lalo CR100% (1)
- TEMA1 Biología Molecular PDFDocument35 pagesTEMA1 Biología Molecular PDFRuben PeñalverNo ratings yet
- Cromosomas humanos y cromosomopatíasDocument27 pagesCromosomas humanos y cromosomopatíasYamir Huarancca ÑaupariNo ratings yet
- Programa GENÉTICA UCA 2022-02Document6 pagesPrograma GENÉTICA UCA 2022-02Rubi Esmeralda Romero SánchezNo ratings yet
- CROMOSOMOPATIASDocument41 pagesCROMOSOMOPATIASEdsonVictorPalomino100% (2)
- 4c-Anomalias CromosomicasDocument5 pages4c-Anomalias CromosomicasManuel HernándezNo ratings yet
- La importancia de la citogenética en el diagnóstico médicoDocument8 pagesLa importancia de la citogenética en el diagnóstico médicoEvert BonifacioNo ratings yet
- Clase Alteraciones CromosomicasDocument57 pagesClase Alteraciones Cromosomicaselfaiteacdg75100% (1)
- Practica 9.-El Cariotipo HumanoDocument4 pagesPractica 9.-El Cariotipo HumanoEmily Diaz VelasquezNo ratings yet
- Citogenetica y El FitomejoramientoDocument7 pagesCitogenetica y El FitomejoramientoEuler Jhonatan Córdova RojasNo ratings yet
- Practica 5 Ideograma HumanoDocument7 pagesPractica 5 Ideograma HumanoAna Maria Niño PerezNo ratings yet
- Alteraciones cromosómicas y síndromes genéticosDocument13 pagesAlteraciones cromosómicas y síndromes genéticosOscar PerezNo ratings yet
- Aneuploidías y EuploidíasDocument11 pagesAneuploidías y EuploidíasMassiel Valenzuela100% (1)
- Trisomia PDFDocument1 pageTrisomia PDFCruz Oteiza Patricia AlejandraNo ratings yet