You might also like
- JonctiuniDocument37 pagesJonctiuniAd AdrianaNo ratings yet
- Curs 1 Tesutul EpitelialDocument11 pagesCurs 1 Tesutul EpitelialVerginia ShuganiNo ratings yet
- Histologie SubiecteDocument18 pagesHistologie SubiecteCosmin NgNo ratings yet
- Membrana CelularaDocument6 pagesMembrana Celularaveronica.99No ratings yet
- Celula Şi Mediul InternDocument225 pagesCelula Şi Mediul InternFlavian Manolache100% (1)
- Prezentare AnticorpiDocument15 pagesPrezentare AnticorpiOanaDorofteNo ratings yet
- PinocitozaDocument26 pagesPinocitozaMonicaNo ratings yet
- Citologie AnimalaDocument28 pagesCitologie AnimalaEmilia Sabau100% (1)
- Curs 01 - 01 - Curs Tesut Epitelial1Document75 pagesCurs 01 - 01 - Curs Tesut Epitelial1Bubulac AngelaNo ratings yet
- C07 CitoscheletulDocument28 pagesC07 Citoscheletulpanterablo100% (1)
- Subiecte Rezolvate ColocviuDocument18 pagesSubiecte Rezolvate ColocviuCristina IoanaNo ratings yet
- Ed. FizDocument2 pagesEd. FizIuliaNo ratings yet
- Citoscheletul PDFDocument41 pagesCitoscheletul PDFOzana CopyShopNo ratings yet
- Citoscheletul MembranarDocument1 pageCitoscheletul MembranarCristina PopNo ratings yet
- Macromolecule ADN ARN Proteine 2015Document36 pagesMacromolecule ADN ARN Proteine 2015Gicu Butuc0% (1)
- Structura ProteinelorDocument13 pagesStructura ProteinelorA B100% (1)
- Manual HistologieDocument93 pagesManual HistologieValiNo ratings yet
- Histologie 1 Curs 3) Tesuturi ConjunctiveDocument7 pagesHistologie 1 Curs 3) Tesuturi ConjunctiveMariaMarya100% (2)
- Lipoproteine PlasmaticeDocument13 pagesLipoproteine PlasmaticeipaperrrNo ratings yet
- Citologie MembranaDocument33 pagesCitologie MembranaOsman Daniela100% (1)
- Mutatii Genice 2012Document43 pagesMutatii Genice 2012Natalia VNo ratings yet
- Organizarea Structurala Si Functionala A CeluleiDocument19 pagesOrganizarea Structurala Si Functionala A CeluleiGiuliaGoth00No ratings yet
- Teste AnatomieDocument43 pagesTeste AnatomiemariutzaNo ratings yet
- Genetica CursDocument4 pagesGenetica CursRonaldo LazarNo ratings yet
- Metabolism Lipidic MDDocument110 pagesMetabolism Lipidic MDElena CristinaNo ratings yet
- Histologie RepereDocument6 pagesHistologie RepereMihaela CiliutaNo ratings yet
- Histologia Generală Şi HistogenezaDocument457 pagesHistologia Generală Şi HistogenezaRaluca MihalacheNo ratings yet
- Secretia GastricaDocument40 pagesSecretia GastricaMadalina LunguNo ratings yet
- Nervii FesieriDocument3 pagesNervii FesieriDan100% (1)
- LimfociteleDocument12 pagesLimfociteleripjojonNo ratings yet
- Reglarea SecretiilorDocument4 pagesReglarea SecretiilorMadalina Mardare0% (1)
- MedicinaDocument421 pagesMedicinaborisNo ratings yet
- Totalizare Anatomie 3Document17 pagesTotalizare Anatomie 3Anonymous ck0aGAqZl100% (2)
- Ciclul ATPDocument12 pagesCiclul ATPAlice Douglas100% (1)
- Biocel LPDocument20 pagesBiocel LPnomatter221No ratings yet
- Intestinul SubtireDocument3 pagesIntestinul SubtirevasilepopNo ratings yet
- Subiecte Exam - An II - Semi Rezolvate.Document80 pagesSubiecte Exam - An II - Semi Rezolvate.MonicaElenaNo ratings yet
- AQUAPORINEDocument26 pagesAQUAPORINEProca IonNo ratings yet
- Examen Biologie CelularaDocument8 pagesExamen Biologie CelularaElena IscuNo ratings yet
- Criterii de Notare Cls A V-ADocument3 pagesCriterii de Notare Cls A V-AAlinaNo ratings yet
- Fiziologia Tesuturilor ExcitabileDocument72 pagesFiziologia Tesuturilor ExcitabileNellu CebotariNo ratings yet
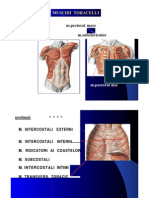
- Muschii ToraceluiDocument7 pagesMuschii ToraceluiBenny ScorpyoNo ratings yet
- Curs VizualDocument31 pagesCurs VizualLarisa P. OanceaNo ratings yet
- Membrana CelularaDocument77 pagesMembrana CelularaCătălina RaicuNo ratings yet
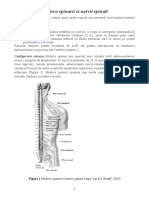
- Maduva Spinarii Si Nervii SpinaliDocument9 pagesMaduva Spinarii Si Nervii SpinalinetprintNo ratings yet
- Curs CitoscheletDocument12 pagesCurs CitoscheletVladDanilaNo ratings yet
- Biochimie SintezeDocument27 pagesBiochimie SintezeChristian DanielNo ratings yet
- DOP. C KrebsDocument7 pagesDOP. C KrebsDaniela LeonteNo ratings yet
- Grile FiziologieDocument20 pagesGrile FiziologieGabriel RusNo ratings yet
- GlucideleDocument7 pagesGlucideleAlexandra MagaleasNo ratings yet
- Curs 3 Aparatul GolgiDocument4 pagesCurs 3 Aparatul GolgiOana Koppandi100% (1)
- Histologie ExamenDocument25 pagesHistologie ExamenMaria Teodora Catană100% (1)
- Curs Histologie Polaritatea Membranei CelulareDocument12 pagesCurs Histologie Polaritatea Membranei CelulareRonaldo LazarNo ratings yet
- Esuturi Excitabile 2022-58228Document131 pagesEsuturi Excitabile 2022-58228Viorica DandeșNo ratings yet
- Subiecte Biologie CelularaDocument11 pagesSubiecte Biologie CelularaBradeanu AndreiNo ratings yet
- Tesut Exitabil 2022-75967Document120 pagesTesut Exitabil 2022-75967Vlad CorciuNo ratings yet
- Tesut Exitabil 2020-16957Document115 pagesTesut Exitabil 2020-16957Bunici MarinelaNo ratings yet
- Biocel Sub ExamDocument12 pagesBiocel Sub ExamAlexandra ConstantinescuNo ratings yet
- Lp4-Specializarile Plasmalemei PDFDocument5 pagesLp4-Specializarile Plasmalemei PDFOvidiu MarianNo ratings yet
- Biologie Celulara - Adeziunea CelularaDocument9 pagesBiologie Celulara - Adeziunea CelularaLaura Pana100% (1)
- Tesutul NervosDocument6 pagesTesutul NervosAriela Săvuţ80% (5)
- Tesutul OsosDocument10 pagesTesutul OsosAriela Săvuţ100% (5)
- Tubul DigestivDocument14 pagesTubul DigestivAriela Săvuţ75% (4)
- Histo 14Document7 pagesHisto 14Alex NicolaeNo ratings yet
- Tesutul ConjunctivDocument7 pagesTesutul ConjunctivAriela SăvuţNo ratings yet
- Tesutul EpitelialDocument6 pagesTesutul EpitelialAriela Săvuţ100% (7)
- Tesutul MuscularDocument4 pagesTesutul MuscularAriela Săvuţ100% (1)
- Sistemul VascularDocument6 pagesSistemul VascularAriela SăvuţNo ratings yet
- HEMATOPOIEZADocument7 pagesHEMATOPOIEZAAriela SăvuţNo ratings yet
- Organele LimfaticeDocument5 pagesOrganele LimfaticeAriela SăvuţNo ratings yet
- Procesul de CheratinizareDocument7 pagesProcesul de CheratinizareAriela SăvuţNo ratings yet
- LeucociteleDocument6 pagesLeucociteleAriela SăvuţNo ratings yet
- Glandele AnexeDocument7 pagesGlandele AnexeAriela SăvuţNo ratings yet
- SANGELEDocument5 pagesSANGELEAriela Săvuţ100% (1)
- OvarulDocument8 pagesOvarulana_maria_neagu_3No ratings yet
- Glande EndocrineDocument7 pagesGlande EndocrineAriela SăvuţNo ratings yet
- Histologie Curs Sem 2Document61 pagesHistologie Curs Sem 2Ariela Săvuţ100% (4)
- StaphylococcusDocument9 pagesStaphylococcusAriela SăvuţNo ratings yet
- Celulele ConjunctiveDocument9 pagesCelulele ConjunctiveAriela SăvuţNo ratings yet
- Aparatul RespiratorDocument6 pagesAparatul RespiratorAriela SăvuţNo ratings yet
- Bacterii AnaerobeDocument6 pagesBacterii AnaerobeAriela Săvuţ100% (1)
- Aparatul Genital FemininDocument9 pagesAparatul Genital FemininAriela Săvuţ100% (2)
- EmbriologieDocument20 pagesEmbriologieAriela Săvuţ100% (1)
- Glanda Pineala (Epifiza)Document7 pagesGlanda Pineala (Epifiza)Ariela Săvuţ100% (2)
- Aparatul Genital MasculinDocument6 pagesAparatul Genital MasculinAriela SăvuţNo ratings yet
- Histologie Curs Sem 2Document61 pagesHistologie Curs Sem 2Ariela Săvuţ100% (4)
- GonococDocument8 pagesGonococAriela Săvuţ100% (1)
- MeningococDocument4 pagesMeningococAriela SăvuţNo ratings yet
- StreptococcusDocument11 pagesStreptococcusAriela Săvuţ100% (5)