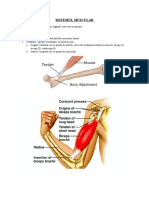
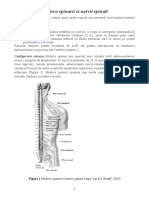
You might also like
- Ghid Pentru Redactarea Si Prezentarea Lucrarii de Licenta, UMFCDDocument6 pagesGhid Pentru Redactarea Si Prezentarea Lucrarii de Licenta, UMFCDghita andreea100% (9)
- Muschiul NetedDocument8 pagesMuschiul NetedAlina Allee100% (1)
- Cursuri NeurologieDocument78 pagesCursuri NeurologieMarinela Cristică100% (1)
- #Micro Sem 1 Subiecte RezolvateDocument34 pages#Micro Sem 1 Subiecte RezolvateRadu DimulescuNo ratings yet
- Silabus Anatomie FunctionalaDocument9 pagesSilabus Anatomie FunctionalaBacso Zsuzsanna100% (1)
- Sindroame HemoragiceDocument93 pagesSindroame HemoragiceAde Byby100% (1)
- Tesutul MuscularDocument8 pagesTesutul MusculardanNo ratings yet
- Anatomia Si Fiziologia OmuluiDocument4 pagesAnatomia Si Fiziologia OmuluiCatargiu AmbrozieNo ratings yet
- Tesuturile AnatomieDocument13 pagesTesuturile AnatomieDan marinescuNo ratings yet
- Subiectele Pentru Examenul de FiziologieDocument6 pagesSubiectele Pentru Examenul de FiziologieAndra CaptanNo ratings yet
- Prezentare ICTERDocument46 pagesPrezentare ICTERpetrys91No ratings yet
- Subiecte Examen Anatomie An I Sem II 2014-2015 FinalDocument3 pagesSubiecte Examen Anatomie An I Sem II 2014-2015 FinalhospitalbucharestNo ratings yet
- Curs 2 Curs 2 Sistemul NervosDocument6 pagesCurs 2 Curs 2 Sistemul NervosIulian Catalin GheorghiuNo ratings yet
- Sistem Vascular - 2018Document10 pagesSistem Vascular - 2018rodicaNo ratings yet
- Scheletul Membrului InferiorDocument25 pagesScheletul Membrului InferiorBalta NicoletaNo ratings yet
- Sistemul PiramidalDocument53 pagesSistemul PiramidalMadalina CordonasuNo ratings yet
- Homeostazia Mediului InternDocument8 pagesHomeostazia Mediului InternMarcu DanielaNo ratings yet
- Sistemul Osos La Om.Document11 pagesSistemul Osos La Om.damian bocanceaNo ratings yet
- Coloana Lombara Curs 8Document6 pagesColoana Lombara Curs 8Gligan Alin-CristianNo ratings yet
- Curs Fiziologie 1Document579 pagesCurs Fiziologie 1Iulian BendreaNo ratings yet
- Osteologie GeneralaDocument52 pagesOsteologie GeneralareeredNo ratings yet
- Anatomie Toate CursurileDocument473 pagesAnatomie Toate CursurileGHIBERDICU IONUT FLORINNo ratings yet
- LeucociteleDocument6 pagesLeucociteleAriela SăvuţNo ratings yet
- Sist Muscular - Cls A XI-aDocument5 pagesSist Muscular - Cls A XI-aUnicorn.100% (1)
- Aparat NeuromuscularDocument26 pagesAparat NeuromuscularAlina-Gabriela Marin50% (2)
- Metabolismul MuscularDocument9 pagesMetabolismul MuscularAndreea Csatlos100% (1)
- Sist - Muscular 2014Document18 pagesSist - Muscular 2014Ioana Coculescu100% (1)
- Reflex eDocument10 pagesReflex eMadalina MarinNo ratings yet
- Tesut Muscular Amalinei PDFDocument16 pagesTesut Muscular Amalinei PDFbDashNo ratings yet
- Maduva SpinariiDocument6 pagesMaduva Spinariicornelia89No ratings yet
- NeuronulDocument61 pagesNeuronulAndreea Radulescu100% (1)
- CelulaDocument12 pagesCelulaIustinian LesinschiNo ratings yet
- Articulatiile CostoDocument1 pageArticulatiile CostoCebotari IvanNo ratings yet
- Curs 1 Biologie Celulara Si MolecularaDocument10 pagesCurs 1 Biologie Celulara Si MolecularaatezorNo ratings yet
- Maduva Spinarii Si Nervii SpinaliDocument9 pagesMaduva Spinarii Si Nervii SpinalinetprintNo ratings yet
- Semio 10Document19 pagesSemio 10Lauren GrayNo ratings yet
- Tesuturile EpitelialeDocument8 pagesTesuturile EpitelialeAlex Seicean100% (1)
- Analizatori UrecheaDocument9 pagesAnalizatori UrecheaIoana PloconNo ratings yet
- Anatomia LocomotorDocument5 pagesAnatomia LocomotorradumariaanaNo ratings yet
- Cursuri An IIDocument93 pagesCursuri An IIVioletaGabrielaIacobNo ratings yet
- Curs Org Si Form LimfoDocument31 pagesCurs Org Si Form LimfoToma CIprian-GeorgeNo ratings yet
- Ecotoxicologie 1Document25 pagesEcotoxicologie 1Ilona RaduNo ratings yet
- Muschiul DeltoidDocument2 pagesMuschiul DeltoidIsabelle CălinNo ratings yet
- M Quadratus LomborumDocument3 pagesM Quadratus LomborumvanciocNo ratings yet
- UlnaDocument2 pagesUlnaRoxana OanceaNo ratings yet
- Curs 4 - Articulatii, PlexDocument7 pagesCurs 4 - Articulatii, PlexCristina IoanaNo ratings yet
- Țesuturile Musculare Sunt Adaptate Funcției de ContracțieDocument3 pagesȚesuturile Musculare Sunt Adaptate Funcției de ContracțieAndriana PrisacariNo ratings yet
- Analizatorul KinestezicDocument25 pagesAnalizatorul KinestezicEne Violeta Alina100% (1)
- Orbita Referat OftalmogieDocument7 pagesOrbita Referat OftalmogieVadim ViţuNo ratings yet
- Sistemul MuscularDocument34 pagesSistemul MuscularIleana AndreeaNo ratings yet
- Oasele Scheletului UmanDocument1 pageOasele Scheletului UmanDiana CristianNo ratings yet
- Biofizica Aparatului LocomotorDocument15 pagesBiofizica Aparatului Locomotorandreeamihai92No ratings yet
- Aparatul CardiovascularDocument8 pagesAparatul CardiovascularAnne BarbuNo ratings yet
- C3 ȚesuturiDocument71 pagesC3 ȚesuturiFlorentina TurtureanNo ratings yet
- Analizatorul CutanatDocument2 pagesAnalizatorul CutanatunafantasiaNo ratings yet
- Analizatorul AuditivDocument2 pagesAnalizatorul AuditivAndrei CaraşNo ratings yet
- Analizatorul CutanatDocument16 pagesAnalizatorul Cutanatpaun_alexandra89No ratings yet
- Functiile Maduvei, Functiile NeuronuluiDocument1 pageFunctiile Maduvei, Functiile NeuronuluilavybbyNo ratings yet
- Sistemul OsosDocument28 pagesSistemul OsosNathan Santiago100% (1)
- Referat1 CoxalDocument3 pagesReferat1 CoxalannalisaizaNo ratings yet
- MuschiiDocument4 pagesMuschiiGeorgePanaitNo ratings yet
- Curs II - Muschiul NetedDocument8 pagesCurs II - Muschiul NetedAlexandra MincaNo ratings yet
- Muschi NetedDocument6 pagesMuschi Netediuuu3No ratings yet
- 5 - MuscularDocument14 pages5 - MuscularVioleta GeorgescuNo ratings yet
- Cursuri Histologie 10,11 An 2, Umf Carol Davilatesutul Muscular, Doamna Doctor Laura CeafalanDocument11 pagesCursuri Histologie 10,11 An 2, Umf Carol Davilatesutul Muscular, Doamna Doctor Laura CeafalanbociocroxanaelenaNo ratings yet
- Special ItDocument6 pagesSpecial Itanomis2999No ratings yet
- Sindromul HipotonDocument89 pagesSindromul HipotonLidia LadeaNo ratings yet
- Studentiepi2 NeuropediatrieDocument32 pagesStudentiepi2 NeuropediatrieLoredana ElenaNo ratings yet
- C3 CerebelDocument42 pagesC3 CerebelSimona BalasescuNo ratings yet
- Lista Examen Urologie FundeniDocument4 pagesLista Examen Urologie FundeniNastasa Petra VandaNo ratings yet
- LesDocument51 pagesLescasimedNo ratings yet
- c19 Bda Si Sda CorectatDocument83 pagesc19 Bda Si Sda CorectatSimona BalasescuNo ratings yet
- Alimentatia Copilului 4Document40 pagesAlimentatia Copilului 4Cristina RaduNo ratings yet
- c1 - Ex Neurologic Prim CursDocument85 pagesc1 - Ex Neurologic Prim CursSimona BalasescuNo ratings yet
- 29 - HIV ScurtDocument39 pages29 - HIV ScurtAde BybyNo ratings yet
- B.patologia AdolescentuluiDocument31 pagesB.patologia AdolescentuluiMihai Bogdan CirsteaNo ratings yet
- Insuf Cardiaca 1Document62 pagesInsuf Cardiaca 1Ade BybyNo ratings yet
- 28 - Deficit Imun CursDocument41 pages28 - Deficit Imun CursAde BybyNo ratings yet
- Notiuni de Baza MGDocument1 pageNotiuni de Baza MGTololoanca ElianNo ratings yet
- HIV La CopilDocument10 pagesHIV La CopilSimona BalasescuNo ratings yet
- Hepatitele Cronice PediatriceDocument68 pagesHepatitele Cronice PediatriceayannapuiNo ratings yet
- AlergeniDocument9 pagesAlergeniSimona BalasescuNo ratings yet
- LP 6 Caractere de CulturaDocument5 pagesLP 6 Caractere de CulturaSimona BalasescuNo ratings yet
- Studentiepi2 NeuropediatrieDocument32 pagesStudentiepi2 NeuropediatrieLoredana ElenaNo ratings yet
- Chestionar RezidentiDocument2 pagesChestionar RezidentiSimona BalasescuNo ratings yet
- EpilepsieDocument3 pagesEpilepsieTerrance RivasNo ratings yet
- GlosarDocument78 pagesGlosarsunshyne_27_mihaNo ratings yet
- Curs Structura AdnDocument44 pagesCurs Structura AdnSimona BalasescuNo ratings yet
- Varietati ConjunctiveDocument2 pagesVarietati ConjunctiveSimona BalasescuNo ratings yet