You might also like
- Figure 2: Hydrolysis of Sucrose (Averill & Eldredge, 2013)Document5 pagesFigure 2: Hydrolysis of Sucrose (Averill & Eldredge, 2013)Hafiidz MalekNo ratings yet
- Nanoemulsion Formulation of Palm Oil Esters ForDocument14 pagesNanoemulsion Formulation of Palm Oil Esters ForVenu Gopal NNo ratings yet
- Biocatalysts and Enzyme TechnologyDocument9 pagesBiocatalysts and Enzyme TechnologyMichelle Santillán0% (1)
- Optimization of Extraction Conditions and Development of A SensitiveDocument6 pagesOptimization of Extraction Conditions and Development of A SensitiveSesilia Bernadet Soubirous MarshellaNo ratings yet
- Enol N Ion EnolatDocument39 pagesEnol N Ion EnolatJulia RahayuNo ratings yet
- Synthesis of AspirinDocument3 pagesSynthesis of AspirinDella Karissa100% (1)
- Class Notes Heterocyclic ChemDocument218 pagesClass Notes Heterocyclic ChemDeepanshu 1459No ratings yet
- (Cis Cu (Gly) 2) H2ODocument13 pages(Cis Cu (Gly) 2) H2OMichaelNo ratings yet
- Efficient Reduction of Imines to Secondary AminesDocument4 pagesEfficient Reduction of Imines to Secondary AminesRatna Siti KhodijahNo ratings yet
- Extraction of Caffeine from Tea LeavesDocument4 pagesExtraction of Caffeine from Tea LeavesTommy Bety100% (1)
- Enzyme PectinaseDocument12 pagesEnzyme PectinaseửeNo ratings yet
- Bioethanol Production From Banana Peels: June 2016Document8 pagesBioethanol Production From Banana Peels: June 2016Charlie Micah AquinoNo ratings yet
- Lab ManualDocument19 pagesLab Manualanon_467104036No ratings yet
- Biosepartaion Engineering: Ch.1: Bioseparation & Biological MaterialsDocument89 pagesBiosepartaion Engineering: Ch.1: Bioseparation & Biological MaterialsAlex MaximusNo ratings yet
- Fragmentation Patterns in The Mass Spectra of Organic CompoundsDocument54 pagesFragmentation Patterns in The Mass Spectra of Organic CompoundsChandra Reddy100% (2)
- The Influence of Conformational Isomerism On Drug ActionDocument6 pagesThe Influence of Conformational Isomerism On Drug Actionhectorlope45No ratings yet
- CatalysisDocument6 pagesCatalysisNaveen PalaNo ratings yet
- Nzymes: By: Mrs. Kalaivani Sathish. M. Pharm, Assistant Professor, Pims - PanipatDocument63 pagesNzymes: By: Mrs. Kalaivani Sathish. M. Pharm, Assistant Professor, Pims - Panipaturmila pandeyNo ratings yet
- Protein Extraction and Characterization TechniquesDocument20 pagesProtein Extraction and Characterization TechniquesMichelle YapNo ratings yet
- Exp7. Isolation of ProteinsDocument6 pagesExp7. Isolation of ProteinsAbdallah BalbaisiNo ratings yet
- Isolation Purification and Identification of CurcuminoidsDocument5 pagesIsolation Purification and Identification of CurcuminoidsNguyenVan HanNo ratings yet
- Prac 4 ReportDocument15 pagesPrac 4 ReportAvanthi Moodley100% (1)
- Determining Lactose in Milk by Refractometry and PolarimetryDocument4 pagesDetermining Lactose in Milk by Refractometry and PolarimetryGabriel Ramirez100% (1)
- A. Heme Pigments: Natural Food Pigments Part 1Document20 pagesA. Heme Pigments: Natural Food Pigments Part 1Balakumaran MNo ratings yet
- 08 Monoglyceride eDocument14 pages08 Monoglyceride eAlf FloNo ratings yet
- Nutritional Analysis of Mungbean CultivarsDocument8 pagesNutritional Analysis of Mungbean CultivarsEuler MiquilenaNo ratings yet
- Proximate Analysis Breakdown of Nesvita Cereal Milk DrinkDocument42 pagesProximate Analysis Breakdown of Nesvita Cereal Milk Drinkfatehatun noorNo ratings yet
- Mass Production of MicroalgaeDocument27 pagesMass Production of Microalgaerajiv pathakNo ratings yet
- 8 Electrozi Ion SelectiviDocument8 pages8 Electrozi Ion SelectiviOana Nicoleta LeoveanuNo ratings yet
- Acetylation of ferrocene experimentDocument5 pagesAcetylation of ferrocene experimentvaibhav palNo ratings yet
- Specification Specification Specification Specification //// Tds TDS TDS TDSDocument2 pagesSpecification Specification Specification Specification //// Tds TDS TDS TDSChâuNo ratings yet
- Extraction and Analysis of LipidsDocument22 pagesExtraction and Analysis of LipidsPauline AlfonsoNo ratings yet
- Ex 6 - TLCDocument9 pagesEx 6 - TLCMonica Hitomi MekaruNo ratings yet
- 11 - Drug Design - Optimizing Target InteractionsDocument67 pages11 - Drug Design - Optimizing Target InteractionsNguyenHongThamNo ratings yet
- What Are HydrogelsDocument4 pagesWhat Are HydrogelsHossein NajafzadehNo ratings yet
- Biol 309 Notes Fall 2016 Final Aug 22Document191 pagesBiol 309 Notes Fall 2016 Final Aug 22Bobby BobertsonNo ratings yet
- Fermentation Technology Chapter I II III IVDocument42 pagesFermentation Technology Chapter I II III IVPuvenez TamalantanNo ratings yet
- Industrial Report On Carew and CompanyDocument60 pagesIndustrial Report On Carew and CompanyShariful IslamNo ratings yet
- Enzyme InformationDocument7 pagesEnzyme InformationLong ThuậnNo ratings yet
- Composition of Milk PDFDocument5 pagesComposition of Milk PDFghanashyam tomarNo ratings yet
- 2-14 Epoxidation of AlkenesDocument2 pages2-14 Epoxidation of AlkenesMohd Zulhelmi AzmiNo ratings yet
- Oxidation ReactionDocument32 pagesOxidation ReactionDeepak PradhanNo ratings yet
- Emulsion Polymerization of Acrylonitrile PDFDocument4 pagesEmulsion Polymerization of Acrylonitrile PDFWalid AdnanNo ratings yet
- Continuous Fermentation of Sago Starch to Solvent Using C. saccharobutylicumDocument9 pagesContinuous Fermentation of Sago Starch to Solvent Using C. saccharobutylicumputrianneNo ratings yet
- Experiments 3 Stage 2017-2018Document50 pagesExperiments 3 Stage 2017-2018Parawgay Danar100% (1)
- Most Effective Lipid Extraction Method from MicroalgaeDocument3 pagesMost Effective Lipid Extraction Method from MicroalgaeRahul GogiaNo ratings yet
- Determination of Water Hardness using Atomic Absorption Spectroscopy (AASDocument7 pagesDetermination of Water Hardness using Atomic Absorption Spectroscopy (AASAfiqah SamanNo ratings yet
- 010 Introduction To Natural Products ChemistryDocument48 pages010 Introduction To Natural Products ChemistryAsnawati AstiNo ratings yet
- DPPH AssayDocument1 pageDPPH AssaySangram DhumalNo ratings yet
- Kgs KGS: Gujarat Alkalies and Chemicals LimitedDocument1 pageKgs KGS: Gujarat Alkalies and Chemicals LimitedChetan Solanki100% (1)
- Total Antioxidant Power and Free RadicalDocument4 pagesTotal Antioxidant Power and Free RadicalDr. Varaprasad Bobbarala100% (1)
- 1 s2.0 S2090123213000969 MainDocument17 pages1 s2.0 S2090123213000969 Mainmarius_alex_32No ratings yet
- Ionic Liquids in Lipid Processing and Analysis: Opportunities and ChallengesFrom EverandIonic Liquids in Lipid Processing and Analysis: Opportunities and ChallengesXuebing XuNo ratings yet
- Transition Metal ToxicityFrom EverandTransition Metal ToxicityG. W. RichterNo ratings yet
- Halogenated Benzenes, Toluenes and Phenols with Water: Solubility Data SeriesFrom EverandHalogenated Benzenes, Toluenes and Phenols with Water: Solubility Data SeriesAri L. HorvathRating: 5 out of 5 stars5/5 (1)
- 993 PushkinDocument0 pages993 PushkinTaher ElBarbaryNo ratings yet
- Brochure ADocument7 pagesBrochure ATaher ElBarbaryNo ratings yet
- Second Wave FeminismDocument12 pagesSecond Wave FeminismTaher ElBarbaryNo ratings yet
- Recognizing Patients High on "Bath SaltsDocument3 pagesRecognizing Patients High on "Bath SaltsTaher ElBarbaryNo ratings yet
- Elementary Math Daily QuestionsDocument217 pagesElementary Math Daily QuestionsTaher ElBarbaryNo ratings yet
- Caffeine Intake, Tolerance, and WithdrawalDocument6 pagesCaffeine Intake, Tolerance, and WithdrawalTaher ElBarbaryNo ratings yet
- Grade 3 Handwriting WorkbookDocument80 pagesGrade 3 Handwriting Workbookpavan_padma977927100% (8)
- Islamic CairoDocument6 pagesIslamic CairoTaher ElBarbaryNo ratings yet
- stb3 Math0606eDocument20 pagesstb3 Math0606eTaher ElBarbaryNo ratings yet
- Association Between A2a Receptor Gene Polymorphisms andDocument9 pagesAssociation Between A2a Receptor Gene Polymorphisms andTaher ElBarbaryNo ratings yet
- Batal Men Hada AlzamanDocument185 pagesBatal Men Hada AlzamanTaher ElBarbaryNo ratings yet
- Democracy Via Translation: Books TitlesDocument5 pagesDemocracy Via Translation: Books TitlesTaher ElBarbaryNo ratings yet
- The Psychology of Atheism: Paul C. VitzDocument12 pagesThe Psychology of Atheism: Paul C. VitzTaher ElBarbaryNo ratings yet
- Animal Models For Anxiety Disorders: Su GuoDocument14 pagesAnimal Models For Anxiety Disorders: Su GuoTaher ElBarbaryNo ratings yet
- WithdrawalDocument5 pagesWithdrawalTaher ElBarbaryNo ratings yet
- The Psychology of Atheism: Paul C. VitzDocument12 pagesThe Psychology of Atheism: Paul C. VitzTaher ElBarbaryNo ratings yet
- Caffeine, Sleep and Wakefulness: Implications of New Under-Standing About Withdrawal ReversalDocument10 pagesCaffeine, Sleep and Wakefulness: Implications of New Under-Standing About Withdrawal ReversalTaher ElBarbaryNo ratings yet
- Association Between A2a Receptor Gene Polymorphisms andDocument9 pagesAssociation Between A2a Receptor Gene Polymorphisms andTaher ElBarbaryNo ratings yet
- EctDocument1 pageEctTaher ElBarbaryNo ratings yet
- Caffeine, A Drug For All SeasonsDocument2 pagesCaffeine, A Drug For All SeasonsTaher ElBarbaryNo ratings yet
- Caffeine Induced PsychosisDocument3 pagesCaffeine Induced PsychosisTaher ElBarbaryNo ratings yet
- Rewriting Cultural Safety Within The.5Document19 pagesRewriting Cultural Safety Within The.5Taher ElBarbaryNo ratings yet
- 8 RecipeForaResearchPaperDocument24 pages8 RecipeForaResearchPaperTaher ElBarbaryNo ratings yet
- 2010 Translation Fund ApplicationDocument2 pages2010 Translation Fund ApplicationTaher ElBarbaryNo ratings yet
- 4Document7 pages4Taher ElBarbaryNo ratings yet
- Anatomy of A Research PaperDocument7 pagesAnatomy of A Research PapermhdstatNo ratings yet
- Ten Steps For Writing Research PapersDocument8 pagesTen Steps For Writing Research PapersSardar AftabNo ratings yet
- Topic 9Document37 pagesTopic 9Taher ElBarbaryNo ratings yet
- Chapter 10Document39 pagesChapter 10Taher ElBarbaryNo ratings yet
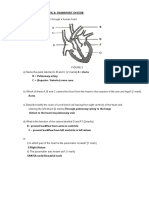
- Transport System Question and AnswersDocument12 pagesTransport System Question and Answerskumara guruparanNo ratings yet
- Final Summative Exam #2 Grade 7Document4 pagesFinal Summative Exam #2 Grade 7Mae CudalNo ratings yet
- T S Form 3Document2 pagesT S Form 3Lubinda Lubinda Jr.No ratings yet
- Effect of pumpkin flour on characteristics of chiffon cake made from modified cassava flourDocument12 pagesEffect of pumpkin flour on characteristics of chiffon cake made from modified cassava flourNurul HasanahNo ratings yet
- 40-SERIES RED POWER COLLECTION BUILT WITH HIGH QUALITY COMPONENTSDocument1 page40-SERIES RED POWER COLLECTION BUILT WITH HIGH QUALITY COMPONENTSB GoodeNo ratings yet
- BandhaDocument3 pagesBandhaMouli ChandraNo ratings yet
- Mech-Design and Fabrication of Motorized Screw Jack For A Four WheelerDocument26 pagesMech-Design and Fabrication of Motorized Screw Jack For A Four WheelerkumarsandiNo ratings yet
- Cp2245 Finish Paint Ea4-5991 MsdsDocument11 pagesCp2245 Finish Paint Ea4-5991 MsdsPanneer SelvamNo ratings yet
- 226-Article Text-515-1-10-20220401Document11 pages226-Article Text-515-1-10-20220401Ann TrầnNo ratings yet
- Climate Factsheets July 2022Document7 pagesClimate Factsheets July 2022Michael SiaNo ratings yet
- Syedna's Zulm On SH Sajjad Husein SarangpuriDocument44 pagesSyedna's Zulm On SH Sajjad Husein SarangpuriMuddasar Ali ZariNo ratings yet
- Product Sheet - MasterFan MF120R ARGB 3in1Document2 pagesProduct Sheet - MasterFan MF120R ARGB 3in1Walter Hugo Friaes PintoNo ratings yet
- Butterfly Valve T-Smart 7 GEADocument2 pagesButterfly Valve T-Smart 7 GEACamilaNo ratings yet
- W-2 Wage and Tax Statement: This Information Is Being Furnished To The Internal Revenue ServiceDocument2 pagesW-2 Wage and Tax Statement: This Information Is Being Furnished To The Internal Revenue ServiceJunk BoxNo ratings yet
- The Marangoni Effect ZulaikhaDocument3 pagesThe Marangoni Effect ZulaikhaZulaikha ZulaikhaNo ratings yet
- HTSX ReportDocument15 pagesHTSX ReportNguyễn Hồng MỹNo ratings yet
- Chemistry ProjectDocument11 pagesChemistry ProjectEktaMallikNo ratings yet
- RD1 Fundic Height MeasurementDocument3 pagesRD1 Fundic Height MeasurementVillanueva JanelleNo ratings yet
- PID Symptoms, Causes, Diagnosis and TreatmentDocument13 pagesPID Symptoms, Causes, Diagnosis and TreatmentAnandila MaulinaNo ratings yet
- Incidence of Candidiasis Amongst Female Students of A Tertiary Institution in Rivers State NigeriaDocument6 pagesIncidence of Candidiasis Amongst Female Students of A Tertiary Institution in Rivers State NigeriaUbali Ibrahim HashimuNo ratings yet
- RVLineDocument28 pagesRVLinesghscribd2012No ratings yet
- Attitudes To Mental Illness 2012 Research Report: Prepared For Time To Change September 2013Document55 pagesAttitudes To Mental Illness 2012 Research Report: Prepared For Time To Change September 2013Jovana AntonijevićNo ratings yet
- Northwest Medicine Travel Screening QuestionnaireDocument2 pagesNorthwest Medicine Travel Screening QuestionnaireMark Philipp AbanNo ratings yet
- Knotless Bankart Repair Using The Labral SwiveLock® and FiberStick™Document6 pagesKnotless Bankart Repair Using The Labral SwiveLock® and FiberStick™savvoNo ratings yet
- Chapter 2 LaboratoryDocument6 pagesChapter 2 LaboratoryVea Roanne GammadNo ratings yet
- A Sonographic Sign of Moderate ToDocument5 pagesA Sonographic Sign of Moderate ToDivisi FER MalangNo ratings yet
- Benign Paroxysmal Migraine Variants of Infancy and Childhood: Transitions and Clinical FeaturesDocument7 pagesBenign Paroxysmal Migraine Variants of Infancy and Childhood: Transitions and Clinical FeaturesNathaly LapoNo ratings yet
- Internship Cafe Coffee Day ReportDocument88 pagesInternship Cafe Coffee Day ReportPoojaNo ratings yet
- 4 Pain Management, Theories of Labor Onset, Signs of LaborDocument31 pages4 Pain Management, Theories of Labor Onset, Signs of Labormamsh KlangNo ratings yet
- Sop Standard Operating ProceduresDocument3 pagesSop Standard Operating ProceduresTaha SaadanNo ratings yet