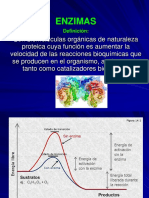
You might also like
- EnzimaDocument13 pagesEnzimaOric Rojas CantuNo ratings yet
- Tema 5. EnzimasDocument10 pagesTema 5. EnzimasPablo VillaescusaNo ratings yet
- .Biologia Comun - Guia #2 de Estudio y Trabajo-4ºmedioDocument9 pages.Biologia Comun - Guia #2 de Estudio y Trabajo-4ºmedioMaribel AlvearNo ratings yet
- Enzimas catalizadoras de la vidaDocument27 pagesEnzimas catalizadoras de la vidaNanci Llampa VilamaniNo ratings yet
- ENZIMASDocument23 pagesENZIMASEsther Salazar Vásquez100% (1)
- Trabajo de EnzimasDocument47 pagesTrabajo de EnzimasMarii Cl ToNo ratings yet
- BIOLOGIA Terminado2Document62 pagesBIOLOGIA Terminado2Milton Cesar Rodrigo TantaleanNo ratings yet
- 34 Article 4Document72 pages34 Article 4Kevin M. MoyaNo ratings yet
- S02A Material S15.s2Document16 pagesS02A Material S15.s2Camila AlessandraNo ratings yet
- Enzimas y catálisisDocument42 pagesEnzimas y catálisissorayakiriachekNo ratings yet
- EnzimasDocument62 pagesEnzimasFaby MoraNo ratings yet
- Clasificación y mecanismos de las enzimasDocument22 pagesClasificación y mecanismos de las enzimasFercho MariscalNo ratings yet
- Introduccion A La EnzimologiaDocument23 pagesIntroduccion A La EnzimologiaFernando FloresNo ratings yet
- Introducción a las enzimas: catalizadores biológicosDocument16 pagesIntroducción a las enzimas: catalizadores biológicosYonahatan QuispeNo ratings yet
- Enzimas: propiedades y especificidadDocument9 pagesEnzimas: propiedades y especificidadMarta Hernández GarcíaNo ratings yet
- Termodinámica y metabolismo celularDocument9 pagesTermodinámica y metabolismo celularmoraNo ratings yet
- Examen 1Document11 pagesExamen 1Alex MejiaNo ratings yet
- B Informe Lab Bioq Amb 2021 2 Practica 6 Actividad Catalasa, Estupiñán Mónica, Peñaranda César, Rojas Wilson, Suarez SaydaDocument16 pagesB Informe Lab Bioq Amb 2021 2 Practica 6 Actividad Catalasa, Estupiñán Mónica, Peñaranda César, Rojas Wilson, Suarez SaydaCESAR DARIO PENARANDA MEDINANo ratings yet
- Informe EnzimasDocument4 pagesInforme EnzimasSindy Alvarez Calle100% (1)
- Qo 9Document24 pagesQo 9JOSE ANTONIO CAYO ABARCANo ratings yet
- Enzimas y VitaminasDocument9 pagesEnzimas y VitaminasRaúlNo ratings yet
- Practica de Accion EnzimaticaDocument8 pagesPractica de Accion EnzimaticaMichaelZhelmiCaEsNo ratings yet
- Enzimas clasificación propiedadesDocument12 pagesEnzimas clasificación propiedadesKiefer MedinaNo ratings yet
- BacteriasDocument4 pagesBacterias5B SyM VARGAS RODRIGUEZ LAURA ALEJANDRANo ratings yet
- Tratamientos EnzimáticosDocument16 pagesTratamientos EnzimáticosJesús Manuel Guerra EstelaNo ratings yet
- Bioquimica Tema PDFDocument20 pagesBioquimica Tema PDFLouise Clemente DoroteoNo ratings yet
- Enzimas Bioq 3Document10 pagesEnzimas Bioq 3Sergio SamitNo ratings yet
- 1° Guía Actividades 8 Metabolismo y EnzimasDocument6 pages1° Guía Actividades 8 Metabolismo y EnzimasYanssuy Lagües PasténNo ratings yet
- Concepto de Enzimas Capsula 1Document4 pagesConcepto de Enzimas Capsula 1Ariel MojicaNo ratings yet
- Ejercicio 1 Bio Quimica Unidad 2Document20 pagesEjercicio 1 Bio Quimica Unidad 2Javier Gonzalez MosqueraNo ratings yet
- Tema5 EnzimasDocument14 pagesTema5 EnzimasIrene PG100% (1)
- Cuestionario Practica EnzimasDocument5 pagesCuestionario Practica EnzimasRocio GutierrezNo ratings yet
- Actividad Enzimática - LaboratorioDocument17 pagesActividad Enzimática - LaboratorioJulio Cesar SamaniegoNo ratings yet
- EnzimasDocument11 pagesEnzimasvanesaNo ratings yet
- Tema 3 - EnzimasDocument3 pagesTema 3 - EnzimasMari Carmen ReinéNo ratings yet
- ENZIMIOLOGÍA - Obstetricia PDFDocument65 pagesENZIMIOLOGÍA - Obstetricia PDFDerecho IcaNo ratings yet
- Practica #2 BioquimicaDocument5 pagesPractica #2 BioquimicaRodoPandaNo ratings yet
- Enzimas 1Document116 pagesEnzimas 1JM JessycaNo ratings yet
- 1610323 B Informe Lab Bioq 2023 1 Práctica 5 Actividad enzimática en la digestiónDocument9 pages1610323 B Informe Lab Bioq 2023 1 Práctica 5 Actividad enzimática en la digestiónYeiner BautistaNo ratings yet
- Enzimas alimentaciónDocument27 pagesEnzimas alimentaciónTOCAS MALCA VICTOR ANDRENo ratings yet
- RESUMENDocument4 pagesRESUMENYirley lucasNo ratings yet
- EnzimasDocument50 pagesEnzimasGato QuequeNo ratings yet
- Biología 3° Curso Plan EspecíficoDocument8 pagesBiología 3° Curso Plan EspecíficoScarlet GrekrovNo ratings yet
- Enzimas: definición, estructura, clasificación y funciónDocument53 pagesEnzimas: definición, estructura, clasificación y funciónNelson BeltranNo ratings yet
- Bioquimica I - Informe 5Document23 pagesBioquimica I - Informe 5Lucero Carhuas IncheNo ratings yet
- Preguntas EnzimologiaDocument2 pagesPreguntas EnzimologiaIVONNE MABELL VEGA NIZANo ratings yet
- Pruebas para La Determinacion de La Velocidad Enzimatica de La CatalasaDocument4 pagesPruebas para La Determinacion de La Velocidad Enzimatica de La CatalasaGiovanni Castillo MaldonadoNo ratings yet
- Tarea No8 BioquimicaDocument6 pagesTarea No8 BioquimicaDaNiel MorenoNo ratings yet
- Tema-7-Biología Cpi-Enzimas y Vitaminas.Document2 pagesTema-7-Biología Cpi-Enzimas y Vitaminas.Christian David Rolón VázquezNo ratings yet
- Enzimas Equipo 5Document9 pagesEnzimas Equipo 5davidNo ratings yet
- Resumen EnzimasDocument9 pagesResumen EnzimasOscarNo ratings yet
- CUESTIONARIODocument11 pagesCUESTIONARIOClaudia PalmaNo ratings yet
- ENZIMASDocument19 pagesENZIMASDaniel LunaNo ratings yet
- Enzimas: Generalidades y características claveDocument29 pagesEnzimas: Generalidades y características claveLuis MarquinezNo ratings yet
- Enzimas y su importancia en las reacciones bioquímicasDocument11 pagesEnzimas y su importancia en las reacciones bioquímicasxiomaGuzmanNo ratings yet
- Taller de EnzimasDocument9 pagesTaller de EnzimasLau Ortiz TerrazaNo ratings yet
- Las Enzimas PDFDocument9 pagesLas Enzimas PDFTriny VilladiegoNo ratings yet
- Cuestionario (Bioquímica)Document13 pagesCuestionario (Bioquímica)rodriguezluism1956No ratings yet
- Decreto 4320Document9 pagesDecreto 4320Leydi Yulieth CastiblancoNo ratings yet
- Cómo Combatir El Pulgón - Cómo Evitarlo y Cómo EliminarloDocument14 pagesCómo Combatir El Pulgón - Cómo Evitarlo y Cómo EliminarloCristhian MorenoNo ratings yet
- Ejercicio 1 de Unidad 3Document11 pagesEjercicio 1 de Unidad 3Cristhian MorenoNo ratings yet
- AccidenteDocument4 pagesAccidenteCristhian MorenoNo ratings yet
- Secado de PapaDocument6 pagesSecado de PapaCristhian MorenoNo ratings yet
- Practica 1 y 2 Li2Document2 pagesPractica 1 y 2 Li2Cristhian MorenoNo ratings yet
- Mapa DestilacionDocument1 pageMapa DestilacionCristhian MorenoNo ratings yet
- GRAFICASDocument1 pageGRAFICASCristhian MorenoNo ratings yet
- Diagram AsDocument3 pagesDiagram AsCristhian Moreno100% (1)
- Triple Boot - Arranque Windows XP, Windows 7 y Windows 8 - Taringa!Document17 pagesTriple Boot - Arranque Windows XP, Windows 7 y Windows 8 - Taringa!Cristhian MorenoNo ratings yet
- Oficio Modificación Responsable SanitarioDocument1 pageOficio Modificación Responsable SanitarioCristhian MorenoNo ratings yet
- Primer Ensayo (Operaciones Unitarias en Ingeníeria Química)Document9 pagesPrimer Ensayo (Operaciones Unitarias en Ingeníeria Química)Pedro DíazNo ratings yet
- Ion Por Arrastre de VaporDocument7 pagesIon Por Arrastre de VaporBrenda SantosNo ratings yet
- 1162511287.clase 3 - Destilación DiferencialDocument11 pages1162511287.clase 3 - Destilación DiferencialEdiriosNo ratings yet
- Destilacion Arrastre de VaporDocument5 pagesDestilacion Arrastre de VaporCristhian Moreno0% (1)
- Simulacion de Proceso de Azucar de CañaDocument3 pagesSimulacion de Proceso de Azucar de CañaCristhian MorenoNo ratings yet
- Diferencia Entre Modelación y Simulación y Los Tipos de SimuladoresDocument15 pagesDiferencia Entre Modelación y Simulación y Los Tipos de SimuladoresCristhian MorenoNo ratings yet
- IQUI-2010-232 Sintesis y Optimizacion de ProcesosDocument8 pagesIQUI-2010-232 Sintesis y Optimizacion de ProcesosAlexis Villalobos JacomeNo ratings yet
- DESTILACIÓNDocument33 pagesDESTILACIÓNCristhian MorenoNo ratings yet
- Análisis y Síntesis de Procesos Ingeniero QuímicoDocument8 pagesAnálisis y Síntesis de Procesos Ingeniero QuímicoCristhian MorenoNo ratings yet
- IQUI-2010-232 Sintesis y Optimizacion de ProcesosDocument8 pagesIQUI-2010-232 Sintesis y Optimizacion de ProcesosAlexis Villalobos JacomeNo ratings yet
- PROTOCOLO (CARBÓN ACTIVADO) WordpadDocument19 pagesPROTOCOLO (CARBÓN ACTIVADO) WordpadCristhian MorenoNo ratings yet
- Y Llegaste TuDocument2 pagesY Llegaste TuCristhian MorenoNo ratings yet
- Cuadro SinopticoDocument1 pageCuadro SinopticoCristhian Moreno0% (1)
- Como Instalar Jelly Bean Por Medio de KDZDocument4 pagesComo Instalar Jelly Bean Por Medio de KDZCristhian MorenoNo ratings yet
- BibliografíaDocument2 pagesBibliografíaCristhian MorenoNo ratings yet
- Bioquimica PDFDocument38 pagesBioquimica PDFCristhian MorenoNo ratings yet
- Actividades de Aprendizaje U3Document3 pagesActividades de Aprendizaje U3Cristhian MorenoNo ratings yet
- Cuestionario de auto-evaluación de electroquímica para ingenieros químicosDocument1 pageCuestionario de auto-evaluación de electroquímica para ingenieros químicosCristhian MorenoNo ratings yet
- ConclusionDocument1 pageConclusionCristhian MorenoNo ratings yet
- Deber Organizador GraficoDocument1 pageDeber Organizador GraficoErikita Valeria LopezNo ratings yet
- Tarea 1Document178 pagesTarea 1Raque PpmNo ratings yet
- Análisis de sistemas hidráulicos laboratorio 02 control electrohidráulico de equipo móvilDocument12 pagesAnálisis de sistemas hidráulicos laboratorio 02 control electrohidráulico de equipo móvilmiguel angel tecse cariNo ratings yet
- Aplico Lo Aprendido Iii B - 6 UnidadDocument3 pagesAplico Lo Aprendido Iii B - 6 UnidadVic EmilioNo ratings yet
- Tarea de Las Placas TectonicasDocument30 pagesTarea de Las Placas TectonicasGerson PerezNo ratings yet
- Guia para Un EiaDocument16 pagesGuia para Un EiaFredy Chumioque UcedaNo ratings yet
- Experiment OsDocument74 pagesExperiment Osdiego santoyo cupiNo ratings yet
- Manejo Conductual en El Aula. XimenaDocument27 pagesManejo Conductual en El Aula. XimenaXimena Covarrubias VarasNo ratings yet
- Fracturas de Pelvis. Presentacion.Document41 pagesFracturas de Pelvis. Presentacion.Maria Fernanda Robles CelisNo ratings yet
- Contaminación y Cambio ClimáticoDocument2 pagesContaminación y Cambio ClimáticoCriis Jurado ArcosNo ratings yet
- 8 Componentes de Una Sonrisa BalanceadaDocument13 pages8 Componentes de Una Sonrisa Balanceadaanthondejesus123No ratings yet
- Ficha de Resumen Modelo Psicodinamico 1.0Document4 pagesFicha de Resumen Modelo Psicodinamico 1.0KAREN XIMENANo ratings yet
- Ficha Ats A4Document8 pagesFicha Ats A4Jorge Luis Espin ClaudioNo ratings yet
- Gritos y Susurros. Introducción A La Inmunorregulación en La Enfermedad PeriodontalDocument6 pagesGritos y Susurros. Introducción A La Inmunorregulación en La Enfermedad PeriodontalFernando Fuentes GonzálezNo ratings yet
- Bibliografia KinesiologiaDocument5 pagesBibliografia KinesiologiaNico AmoresNo ratings yet
- Impacto de la Revolución Verde en la agricultura ecológicaDocument2 pagesImpacto de la Revolución Verde en la agricultura ecológica6621910034 MARIA JOSÉMORELO WILCHES ESTUDIANTE ACTIVONo ratings yet
- 1silices en AguaDocument140 pages1silices en AguaSergio Linares AlonsNo ratings yet
- Eyaculacion Precoz Manual PDFDocument9 pagesEyaculacion Precoz Manual PDFinf196250% (2)
- Test Ion AmonioDocument1 pageTest Ion AmonioAleNo ratings yet
- CH CH (CH) CH CH CH CH: Iso BDocument20 pagesCH CH (CH) CH CH CH CH: Iso BNathalia AcuñaNo ratings yet
- Guiones laboratorio GEO I y IIDocument20 pagesGuiones laboratorio GEO I y IISergi Royo TroNo ratings yet
- Trastorno Bipolar PLACESDocument12 pagesTrastorno Bipolar PLACESDayanna Munguia100% (1)
- I Trim Cta 5ºDocument45 pagesI Trim Cta 5ºRomel Rodrimo RenoguezNo ratings yet
- Presentación de PowerPoint SenaDocument1 pagePresentación de PowerPoint SenaJorge ivan0% (1)
- Resumen Lágrima de BosqueDocument12 pagesResumen Lágrima de BosqueValentina60% (10)
- Tipos de virus informáticos y sus característicasDocument2 pagesTipos de virus informáticos y sus característicassarita torresNo ratings yet
- Taller HABILIDADES SOCIALES INFORMATICA GinaDocument21 pagesTaller HABILIDADES SOCIALES INFORMATICA GinaSorleyda Medina RamirezNo ratings yet
- El Amor de Dios Es JesusDocument33 pagesEl Amor de Dios Es JesusMariano Damian ChiodoNo ratings yet
- 1 Semana - El TextoDocument15 pages1 Semana - El Textorosalina yahuana huamanNo ratings yet
- Capitulo 5Document23 pagesCapitulo 5Roy Umaña CarrilloNo ratings yet