You might also like
- Organs of VisionDocument3 pagesOrgans of VisionNarasimha MurthyNo ratings yet
- Dr. Olasupo S. AdeniyiDocument89 pagesDr. Olasupo S. AdeniyiUloko ChristopherNo ratings yet
- The Eye Pre-Lab: s94 s98 Index of ImagesDocument13 pagesThe Eye Pre-Lab: s94 s98 Index of ImagesAndi KasiantoNo ratings yet
- Anatomy and Function of The EyeDocument3 pagesAnatomy and Function of The EyeAljhon DelfinNo ratings yet
- 15making Sense of ItDocument12 pages15making Sense of Itapi-159564163No ratings yet
- Anatomy of The EYEDocument117 pagesAnatomy of The EYELancelot BritaniaNo ratings yet
- The Anatomy and Physiology of The Eye 3 Classes Objects 1. Master The Anatomy and Physiology of The Eye 2. Relationship With Clinical DiseasesDocument181 pagesThe Anatomy and Physiology of The Eye 3 Classes Objects 1. Master The Anatomy and Physiology of The Eye 2. Relationship With Clinical DiseasessanjivdasNo ratings yet
- Anatomy and Physiology of The EyesDocument3 pagesAnatomy and Physiology of The EyesNkk Aqnd MgdnglNo ratings yet
- EyeballDocument14 pagesEyeballMirko BelanNo ratings yet
- Vision: Chapter 9: SensesDocument21 pagesVision: Chapter 9: SenseshoneykrizelNo ratings yet
- PBL Eye No1,2,3Document9 pagesPBL Eye No1,2,3adelaNo ratings yet
- Sense Organs: Eyes, Ears, Tongue, Nose & SkinDocument132 pagesSense Organs: Eyes, Ears, Tongue, Nose & SkinChandan ShahNo ratings yet
- Visual Disorder Insights: Altered Perception PatternsDocument7 pagesVisual Disorder Insights: Altered Perception PatternsJobelle AcenaNo ratings yet
- UntitledDocument45 pagesUntitledc/risaaq yuusuf ColoowNo ratings yet
- Everyday Science NotesDocument46 pagesEveryday Science NotesGul JeeNo ratings yet
- The EyeDocument11 pagesThe EyeJaja SolasNo ratings yet
- Topic Presentation CataractDocument11 pagesTopic Presentation Cataractjyoti singhNo ratings yet
- BLUNT TRAUMA TO THE EYEDocument38 pagesBLUNT TRAUMA TO THE EYEAyu KottenNo ratings yet
- Yoga & Eye - Anatomy and PhysiologyDocument73 pagesYoga & Eye - Anatomy and Physiologysachi sharma0% (1)
- Anatomyof EyesDocument7 pagesAnatomyof EyesFair Lemming AdamNo ratings yet
- Anatomy and Physiology of The Human EyeDocument9 pagesAnatomy and Physiology of The Human EyeMaryy Basilio100% (2)
- B & C, Vol-12 PDFDocument4 pagesB & C, Vol-12 PDFKiran RmvNo ratings yet
- Ophthalmology 2Document235 pagesOphthalmology 2yididiyagirma30No ratings yet
- The Best Place for CSS Aspirants to Learn About the Anatomy and Function of the Human EyeDocument50 pagesThe Best Place for CSS Aspirants to Learn About the Anatomy and Function of the Human EyeWaqas Gul100% (1)
- Seminar On GlaucomaDocument26 pagesSeminar On GlaucomaPriya ANo ratings yet
- Histology of Cornea & RetinaDocument14 pagesHistology of Cornea & RetinaSandhya KurupNo ratings yet
- Visual DisordersDocument21 pagesVisual DisordersJobelle Acena100% (1)
- Eye Lecture GuideDocument46 pagesEye Lecture Guidemaj0% (1)
- 1-Anatomy of The Eye BallDocument6 pages1-Anatomy of The Eye BalltressNo ratings yet
- Visual SystemDocument6 pagesVisual SystemJobelle Acena100% (1)
- The eye: organ of sightDocument6 pagesThe eye: organ of sightDebiprasad GhoshNo ratings yet
- Lens (Anatomy) : Biconvex Eye Cornea Refract Light Focused Retina Focal Distance Real Image Accommodation Below LensesDocument11 pagesLens (Anatomy) : Biconvex Eye Cornea Refract Light Focused Retina Focal Distance Real Image Accommodation Below LensesvrlftrNo ratings yet
- Anatomy of The EyeDocument4 pagesAnatomy of The EyeSabrinaAyuPutriNo ratings yet
- Glaucoma 180813151731Document24 pagesGlaucoma 180813151731malathiNo ratings yet
- Info Sheet 4 - 1Document8 pagesInfo Sheet 4 - 1Francis ObmergaNo ratings yet
- Eye Anatomy and Diseases ExplainedDocument6 pagesEye Anatomy and Diseases ExplainedFaizal IkhsanNo ratings yet
- ANATOMY & PHYSIOLOGY OF THE EYEDocument70 pagesANATOMY & PHYSIOLOGY OF THE EYEArif MohammadNo ratings yet
- Anatomy and Physiology of The EyeDocument6 pagesAnatomy and Physiology of The Eyesen_subhasis_58No ratings yet
- Sense OrgansDocument12 pagesSense Organsriwisi2139No ratings yet
- Section 13 - Chapter 6 Anatomy and Structure of The EyeDocument12 pagesSection 13 - Chapter 6 Anatomy and Structure of The EyeMosteanu IulianNo ratings yet
- Eyes Definition: Last Updated: September 7, 2017Document5 pagesEyes Definition: Last Updated: September 7, 2017Aljhon DelfinNo ratings yet
- Technical College of Health ShekhanDocument9 pagesTechnical College of Health ShekhanSaadNo ratings yet
- Anatomy of LensDocument20 pagesAnatomy of LensKina KinaNo ratings yet
- EYESIGHT EXPLAINEDDocument10 pagesEYESIGHT EXPLAINEDromaNo ratings yet
- Special Sense EYEDocument74 pagesSpecial Sense EYEprekshya shresthaNo ratings yet
- Anatomy and Physiology of The EyeDocument22 pagesAnatomy and Physiology of The EyeBalkos 61No ratings yet
- Eye and AdnexaDocument22 pagesEye and AdnexaWadabiNo ratings yet
- Ocular PhysiologyDocument80 pagesOcular PhysiologyKenny KayNo ratings yet
- 5 Senses and Their Sense OrgansDocument21 pages5 Senses and Their Sense OrgansViswaNo ratings yet
- Anatomy of EyesDocument7 pagesAnatomy of EyesFrancesc Richard WengerNo ratings yet
- Human EyeDocument54 pagesHuman EyeRadu VisanNo ratings yet
- (L-2) - Eye - Nov 12, 2019Document54 pages(L-2) - Eye - Nov 12, 2019Sejal PrasadNo ratings yet
- Eye & Ear Embryology ObjectivesDocument3 pagesEye & Ear Embryology ObjectivesDr. DNo ratings yet
- Notes On Eye of RabbitDocument3 pagesNotes On Eye of RabbitShailesh NiroulaNo ratings yet
- Anatomy and Physiology ofDocument4 pagesAnatomy and Physiology ofAyu HartantiNo ratings yet
- Anatomy of The EyeDocument3 pagesAnatomy of The EyeManish SharafNo ratings yet
- Low Vision: Assessment and Educational Needs: A Guide to Teachers and ParentsFrom EverandLow Vision: Assessment and Educational Needs: A Guide to Teachers and ParentsNo ratings yet
- A Simple Guide to the Eye and Its Disorders, Diagnosis, Treatment and Related ConditionsFrom EverandA Simple Guide to the Eye and Its Disorders, Diagnosis, Treatment and Related ConditionsNo ratings yet
- The Insect World: Being a Popular Account of the Orders of Insects; Together with a Description of the Habits and Economy of Some of the Most Interesting SpeciesFrom EverandThe Insect World: Being a Popular Account of the Orders of Insects; Together with a Description of the Habits and Economy of Some of the Most Interesting SpeciesNo ratings yet
- 5 - The Visual System and Mechanisms of PerceptionDocument13 pages5 - The Visual System and Mechanisms of PerceptionElmer SalazarNo ratings yet
- Mata Tenang Visus Turun MendadakDocument74 pagesMata Tenang Visus Turun MendadakYeni AnggrainiNo ratings yet
- BioPsy 5 - SensationsDocument13 pagesBioPsy 5 - SensationsTim AroscoNo ratings yet
- A Textbook of Ophthalmology 2nd Edition - Ahmed - 2001Document535 pagesA Textbook of Ophthalmology 2nd Edition - Ahmed - 2001ximemq100% (1)
- Makalah Miopi Bahasa InggrisDocument16 pagesMakalah Miopi Bahasa InggrisDhewii Amelya MiissLoversNo ratings yet
- CSE Artificial Eye REPORTDocument18 pagesCSE Artificial Eye REPORTVaibhav TiwariNo ratings yet
- Joukal (2017) Anatomy of The Human Visual PathwayDocument16 pagesJoukal (2017) Anatomy of The Human Visual PathwaysektemujibNo ratings yet
- Medicolegal Aspects of DeathDocument26 pagesMedicolegal Aspects of DeathRonald Alasa-as AtigNo ratings yet
- VETERINARY Techniques of Ophthalmic Examination ManualDocument29 pagesVETERINARY Techniques of Ophthalmic Examination ManualLuis Torres AlemarNo ratings yet
- Sensory Systems Kd12Document32 pagesSensory Systems Kd12Diana Quimpan CilloNo ratings yet
- Brain ChipDocument24 pagesBrain ChipVINAYAK CODMNo ratings yet
- Assessing EyesDocument33 pagesAssessing Eyesshellacayetano2No ratings yet
- Palm Vein TechnologyDocument29 pagesPalm Vein TechnologyParvathi GoudNo ratings yet
- Chapter 3 &4 ReviewerDocument18 pagesChapter 3 &4 Reviewersophia sierraNo ratings yet
- GlaucomaDocument23 pagesGlaucomaToshiba Satellite0% (1)
- Special SensesDocument4 pagesSpecial SensesGrayson JamesNo ratings yet
- QP May-2016 Paper 1 PDFDocument32 pagesQP May-2016 Paper 1 PDFMoazzem HossainNo ratings yet
- OMS 800 UmDocument109 pagesOMS 800 UmJavi Payá HerrerosNo ratings yet
- Pathogenic mechanism of eye contusionDocument2 pagesPathogenic mechanism of eye contusionSuzan LaiNo ratings yet
- Information ProcessingDocument51 pagesInformation ProcessingNurizni100% (1)
- Review of Various Biometric Authentication TechniquesDocument7 pagesReview of Various Biometric Authentication TechniquesIJIRAE- International Journal of Innovative Research in Advanced EngineeringNo ratings yet
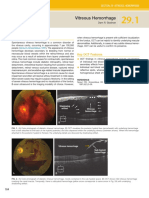
- Vitreous HemorrhageDocument2 pagesVitreous HemorrhageTony Miguel Saba SabaNo ratings yet
- What Is An Epiretinal Membrane?Document6 pagesWhat Is An Epiretinal Membrane?Chip MorrisNo ratings yet
- Gene - and Cell-Based Treatment Strategies For The EyeDocument101 pagesGene - and Cell-Based Treatment Strategies For The EyeCantika MutiaraNo ratings yet
- Classification of Choroidal Neovascularization TypesDocument1 pageClassification of Choroidal Neovascularization TypesJehan Fauzi RNo ratings yet
- Homework #1: Spring 2014 Csce 763: Digital Image ProcessingDocument4 pagesHomework #1: Spring 2014 Csce 763: Digital Image ProcessingNgọc ÁnhNo ratings yet
- S V Engineering College: Lecture NotesDocument123 pagesS V Engineering College: Lecture NotesDeepika AkkiniNo ratings yet
- Optic Disk Hemorrhage in Health and DiseaseDocument19 pagesOptic Disk Hemorrhage in Health and DiseaseTony Miguel Saba SabaNo ratings yet
- Fully Automated Fluorescein AngiographyDocument3 pagesFully Automated Fluorescein AngiographyHamza ElmbarkiNo ratings yet
- DT LeukocoriaDocument44 pagesDT LeukocoriahamzahNo ratings yet