You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
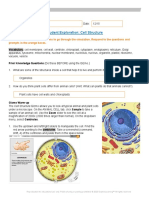
- Cell Structure GizmoDocument4 pagesCell Structure GizmoDavid Yee57% (7)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- 2 The Cell (PDFDrive)Document51 pages2 The Cell (PDFDrive)audiNo ratings yet
- Science: Cell: The Basic Structural and Functional Unit of LifeDocument37 pagesScience: Cell: The Basic Structural and Functional Unit of LifeJaycelyn BaduaNo ratings yet
- Cell Organelles WorksheetDocument8 pagesCell Organelles Worksheete50% (2)
- Cell Organelles and Their FunctionsDocument22 pagesCell Organelles and Their FunctionsJoseph Al Bayonas100% (1)
- Biology Chapter 1 Cell Notes.Document6 pagesBiology Chapter 1 Cell Notes.Jyoti AmbwaniNo ratings yet
- From Tough Nuts To Touch-Me-NotsDocument2 pagesFrom Tough Nuts To Touch-Me-NotsycplainNo ratings yet
- A Molecular Link Between Stem Cell Regulation and Floral Patterning in ArabidopsisDocument11 pagesA Molecular Link Between Stem Cell Regulation and Floral Patterning in ArabidopsisycplainNo ratings yet
- Building Beauty The Genetic Control of Floral PatterningDocument8 pagesBuilding Beauty The Genetic Control of Floral PatterningycplainNo ratings yet
- Building Beauty The Genetic Control of Floral PatterningDocument8 pagesBuilding Beauty The Genetic Control of Floral PatterningycplainNo ratings yet
- A Molecular Link Between Stem Cell Regulation and Floral Patterning in ArabidopsisDocument11 pagesA Molecular Link Between Stem Cell Regulation and Floral Patterning in ArabidopsisycplainNo ratings yet
- Plant Cytokinesis Fission by FusionDocument7 pagesPlant Cytokinesis Fission by FusionycplainNo ratings yet
- Essential Biology 2.2 and 2.3Document7 pagesEssential Biology 2.2 and 2.3MrNewbsNo ratings yet
- Mentoring (Mod 6)Document26 pagesMentoring (Mod 6)Mayrigen DominguezNo ratings yet
- Worksheet 1.1 Cell OrganelleDocument2 pagesWorksheet 1.1 Cell OrganelleCyndel TindoyNo ratings yet
- My Answer in Our LaboratoryDocument4 pagesMy Answer in Our LaboratoryKris Marie DelaCruz DomingoNo ratings yet
- LisosomDocument3 pagesLisosomAnonymous O1xkZINNo ratings yet
- Ieep 105Document7 pagesIeep 105Usha PalNo ratings yet
- M2C The Nucleus QuestionsDocument10 pagesM2C The Nucleus QuestionsJhanna MooreNo ratings yet
- FSC Notes Biology Part 1 Chapter 4 The CellDocument19 pagesFSC Notes Biology Part 1 Chapter 4 The CellGhulam Qasim QaisraniNo ratings yet
- Cell Organelles and Their FunctionsDocument3 pagesCell Organelles and Their FunctionsAly FayeNo ratings yet
- BIO 201 Chapter 3, Part 2 LectureDocument36 pagesBIO 201 Chapter 3, Part 2 LectureDrPearcyNo ratings yet
- Botany Bio Molecules Question Pap WR See TterDocument60 pagesBotany Bio Molecules Question Pap WR See TterSURESH SURESHNo ratings yet
- Rap CellsDocument1 pageRap CellsLowiella MorillaNo ratings yet
- The Cell: The Basic Unit of LifeDocument26 pagesThe Cell: The Basic Unit of Lifeحفيظو عبداللهNo ratings yet
- Intracellular Compartments and Protein SortingDocument9 pagesIntracellular Compartments and Protein SortingJudith UgwujaNo ratings yet
- Cell Division: Mitosis and Cytokinesis: Process Diagrams Step-by-StepDocument7 pagesCell Division: Mitosis and Cytokinesis: Process Diagrams Step-by-Steprambabs369No ratings yet
- Create 3D Cell Model by Deadline for Biology GradeDocument4 pagesCreate 3D Cell Model by Deadline for Biology GradeYohann TuazonNo ratings yet
- Cell Organelle Functions and AnalogiesDocument27 pagesCell Organelle Functions and Analogiesbugboybugboy0% (1)
- Cell Organelles 2Document13 pagesCell Organelles 2Sadeeq ur RahmanNo ratings yet
- The Fundamental Unit of LifeDocument19 pagesThe Fundamental Unit of Lifethinkiit100% (1)
- Nukleus Dan NukleolusDocument63 pagesNukleus Dan NukleolusKarin NakajimaNo ratings yet
- Learn Cell Biology with Fun Songs and Rap LyricsDocument7 pagesLearn Cell Biology with Fun Songs and Rap Lyricsᜊᜎᜑᜎ ᜋᜃᜓᜐᜓᜄ ᜋᜆᜈᜇNo ratings yet
- Cytoplasm (Author V.Mazuru) PDFDocument46 pagesCytoplasm (Author V.Mazuru) PDFLexa KpvNo ratings yet
- Chapter 4 A Tour of The CellDocument12 pagesChapter 4 A Tour of The Cellhuy luongNo ratings yet
- The Fundamental Unit of LifeDocument38 pagesThe Fundamental Unit of LifevpsandhyaraamNo ratings yet