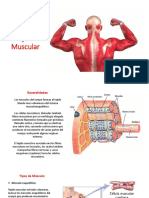
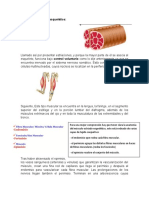
You might also like
- Clase 13 TejidoMuscularDocument30 pagesClase 13 TejidoMuscularSolange Colina0% (1)
- Manual CorrectoDocument41 pagesManual CorrectoCiber Centro100% (1)
- Tejido MuscularDocument20 pagesTejido MuscularSebastián MendezNo ratings yet
- Fisiologia Del Adulto MayorDocument36 pagesFisiologia Del Adulto MayorHenry Trujillo MaytaNo ratings yet
- Articulos de Yoga para Bajar de PesoDocument29 pagesArticulos de Yoga para Bajar de PesoKatherine AbreuNo ratings yet
- Importancia del sistema muscular: sus funciones y cuidadosDocument3 pagesImportancia del sistema muscular: sus funciones y cuidadosmileNo ratings yet
- Proceso de EnvejecimientoDocument18 pagesProceso de EnvejecimientoGianela Noemí Tejada VasquezNo ratings yet
- Empresas, Directivos y Trabajadores ToxicosDocument22 pagesEmpresas, Directivos y Trabajadores ToxicosRafael PazNo ratings yet
- Tejido MuscularDocument31 pagesTejido MuscularLastJd100% (2)
- Músculo LisoDocument3 pagesMúsculo LisoLianGaming HDNo ratings yet
- Tema 19 (Tejido Muscular)Document37 pagesTema 19 (Tejido Muscular)Carlos Daniel Cárdenas SantosNo ratings yet
- MANUAL DE LAB 3 Estudiantes 2023Document11 pagesMANUAL DE LAB 3 Estudiantes 2023Carlos Mauricio VizcarraNo ratings yet
- Tejido muscular: clasificación y funcionesDocument8 pagesTejido muscular: clasificación y funcionesDARWIN RUDY CAMPERO GUZMÁNNo ratings yet
- Información 3er Examen de HistologíaDocument37 pagesInformación 3er Examen de HistologíaPEDRO HONORATO CHAMBA ORDINOLANo ratings yet
- Tejido Muscular HistologíaDocument22 pagesTejido Muscular HistologíaLuiisPascaciioNo ratings yet
- Cuáles Son Los Tipos de Tejido MuscularDocument12 pagesCuáles Son Los Tipos de Tejido Muscularelfisico666No ratings yet
- Trabajo Tejido MuscularDocument10 pagesTrabajo Tejido Muscularelismer gamez100% (1)
- LocomacionDocument7 pagesLocomacionCurso HD USM Sección ANo ratings yet
- Tejido MuscularDocument33 pagesTejido MuscularRocioNo ratings yet
- Funciones músculo esqueléticoDocument17 pagesFunciones músculo esqueléticoKaren GarcíaNo ratings yet
- Tejido MuscularDocument12 pagesTejido MuscularMaria Virginia Alvarado LopezNo ratings yet
- Tejido muscular: características de los tipos esquelético, cardiaco y lisoDocument7 pagesTejido muscular: características de los tipos esquelético, cardiaco y lisoLabquen Kiyen Llanquileo SalazarNo ratings yet
- Tejido MuscularDocument63 pagesTejido MuscularDannoxNo ratings yet
- Musculo Primera ParteDocument21 pagesMusculo Primera ParteCristian OrlegoNo ratings yet
- Tejido MuscularDocument13 pagesTejido MusculardanielaNo ratings yet
- Anatomia y Fisiologia Del Sistema MuscularDocument22 pagesAnatomia y Fisiologia Del Sistema MuscularJ. Gabriel Torres67% (6)
- Tejido MuscularDocument14 pagesTejido MuscularDay BravoNo ratings yet
- Sistema MuscularDocument9 pagesSistema MuscularCarlos GaetanNo ratings yet
- Tipos de Tejido MuscularDocument1 pageTipos de Tejido MuscularjesusNo ratings yet
- Tejido Muscular Material Complementario (Histología)Document12 pagesTejido Muscular Material Complementario (Histología)miguel bravoNo ratings yet
- Guía Sistema MuscularDocument15 pagesGuía Sistema MuscularCHELCY DIAZ GARCESNo ratings yet
- 3 Tejido MuscularDocument9 pages3 Tejido MuscularJemme Carolina López MauriceNo ratings yet
- Tejido Muscular ResumenDocument21 pagesTejido Muscular ResumenEl KurkoNo ratings yet
- Proyecto de AnatomiaDocument15 pagesProyecto de AnatomiaAna PadillaNo ratings yet
- Tejido Muscular (Document29 pagesTejido Muscular (naomi orozcoNo ratings yet
- SISTEMA MUSCULAR- Reposo - © 2003 Edgar Lopategui CorsinoDocument16 pagesSISTEMA MUSCULAR- Reposo - © 2003 Edgar Lopategui CorsinoBAJA DE PESO SIN REBOTENo ratings yet
- Tejidos Cardiaco Estriado y LisoDocument9 pagesTejidos Cardiaco Estriado y LisoDorianaNo ratings yet
- 08 - Sistema MuscularDocument16 pages08 - Sistema Muscularbio.agus92No ratings yet
- Tejido muscular: estructura, tipos y característicasDocument4 pagesTejido muscular: estructura, tipos y característicasLorena MoreiraNo ratings yet
- Tejido muscular: tipos, características y funcionesDocument77 pagesTejido muscular: tipos, características y funcionesGiovanna Vega100% (1)
- Estructra y Funciones Del Muscculo EsqueleticoDocument13 pagesEstructra y Funciones Del Muscculo EsqueleticoMaricruz Cordova DuarteNo ratings yet
- Funcion de Los Musculos SuperficialesDocument56 pagesFuncion de Los Musculos SuperficialesMaria Acosta100% (3)
- El Músculo Estriado EsqueléticoDocument3 pagesEl Músculo Estriado EsqueléticoDaniela CardonaNo ratings yet
- Tejido muscular: estructura y tiposDocument28 pagesTejido muscular: estructura y tiposCHRISTIAN ALBERTO AVILA TORRESNo ratings yet
- Histología Del Sistema MuscularDocument1 pageHistología Del Sistema Muscularvsq6hwm52vNo ratings yet
- Tejido MuscularDocument11 pagesTejido MuscularGraciela Romero PalacioNo ratings yet
- Biology Subject For High School Upper & Lower Arm Muscles by SlidesgoDocument33 pagesBiology Subject For High School Upper & Lower Arm Muscles by Slidesgobykemy54No ratings yet
- Manual Avanzado Capacitación en Elaboración de CárnicosDocument149 pagesManual Avanzado Capacitación en Elaboración de CárnicosRolando GandolfoNo ratings yet
- Tejido MuscularDocument10 pagesTejido MuscularMau MartinezNo ratings yet
- Modulo 1 - Miología GeneralDocument7 pagesModulo 1 - Miología GeneralmartavicNo ratings yet
- Tejido Muscular Estriado EsqueleticoDocument7 pagesTejido Muscular Estriado EsqueleticoMiguel Angel Solorzano ParedesNo ratings yet
- Tema 1. Sistema OsteomusculoarticularDocument63 pagesTema 1. Sistema Osteomusculoarticularjulin400No ratings yet
- Guia de Identificacion Microscópica Del Tejido MuscularDocument4 pagesGuia de Identificacion Microscópica Del Tejido MusculardanielNo ratings yet
- Tejido EpitelialDocument18 pagesTejido EpitelialDavid Laiker MartiinezNo ratings yet
- SISTEMA MUSCULARDocument4 pagesSISTEMA MUSCULARLudmila AcuñaNo ratings yet
- Actividad 7. Sistema MuscularDocument4 pagesActividad 7. Sistema MuscularJorge Luis Merino GuarderasNo ratings yet
- Histologia CuestionarioDocument6 pagesHistologia CuestionarioDayarling ArciaNo ratings yet
- MúsculoDocument77 pagesMúsculoNancy MoranchelNo ratings yet
- TEJIDO MUSCULAR Y ÓSEODocument16 pagesTEJIDO MUSCULAR Y ÓSEORotsen MoralesNo ratings yet
- Informe de Laboratorio 4. Grupo 6Document15 pagesInforme de Laboratorio 4. Grupo 6christian vera0% (1)
- Tejido MuscularDocument12 pagesTejido MuscularEduardo GálvezNo ratings yet
- Mioanatomía animalDocument9 pagesMioanatomía animalDeysi Morales MaqueraNo ratings yet
- Sistema MuscularDocument5 pagesSistema MuscularPauloMazziniNo ratings yet
- Tejido MuscularDocument20 pagesTejido MuscularIdalia GermánNo ratings yet
- Coloquio IXDocument5 pagesColoquio IXRolleros unidosNo ratings yet
- La Salud Como ProcesoDocument1 pageLa Salud Como ProcesoRafael PazNo ratings yet
- Programa de Prevención de Enfermedades Musculoesqueléticas para Las RedesDocument58 pagesPrograma de Prevención de Enfermedades Musculoesqueléticas para Las RedesRafael PazNo ratings yet
- Ejercicios Trabajo ComunitarioDocument1 pageEjercicios Trabajo ComunitarioRafael PazNo ratings yet
- Trabajo Rio 2do RedDocument17 pagesTrabajo Rio 2do RedRafael PazNo ratings yet
- MORFO - Metabolismo de Los LípidosDocument14 pagesMORFO - Metabolismo de Los LípidosRafael PazNo ratings yet
- Copia de Diseño Morfofisiología I - ADIDocument47 pagesCopia de Diseño Morfofisiología I - ADIRafael PazNo ratings yet
- Salud Mental en Atencion PrimariaDocument11 pagesSalud Mental en Atencion PrimariaRafael PazNo ratings yet
- MORFO - Metabolismo de Los LípidosDocument16 pagesMORFO - Metabolismo de Los LípidosRafael PazNo ratings yet
- TEMA 1 Trabajo Rio Primer Semestre RedDocument19 pagesTEMA 1 Trabajo Rio Primer Semestre RedRafael PazNo ratings yet
- Tejido Epitelial..!!!Document66 pagesTejido Epitelial..!!!Rafael Paz100% (1)
- Monografía-Grupo2-Seccion2!Tejido Cartilaginoso y TejidoOseo!2003!Document21 pagesMonografía-Grupo2-Seccion2!Tejido Cartilaginoso y TejidoOseo!2003!Rafael Paz100% (1)
- Embriología-Tejido ConjuntivoDocument4 pagesEmbriología-Tejido ConjuntivoRafael PazNo ratings yet
- Monografia. Metabolismo de Los Carbohidratos Grupo4Document16 pagesMonografia. Metabolismo de Los Carbohidratos Grupo4Rafael PazNo ratings yet
- Metabolismo de Los Carbohidratos. Grupo4!Document15 pagesMetabolismo de Los Carbohidratos. Grupo4!Rafael Paz100% (1)
- Programacion Unefm Medicina SilvaDocument1 pageProgramacion Unefm Medicina SilvaRafael PazNo ratings yet
- Lipido 1Document9 pagesLipido 1Rafael Paz100% (1)
- Lisosoma. Sección2. Grupo4Document6 pagesLisosoma. Sección2. Grupo4Rafael PazNo ratings yet
- EmbriologiaDocument7 pagesEmbriologiaMaria ChirinosNo ratings yet
- Conjuntivo 1Document29 pagesConjuntivo 1Rafael PazNo ratings yet
- Oseo 1Document41 pagesOseo 1Rafael PazNo ratings yet
- Mitocondria 1Document23 pagesMitocondria 1Rafael Paz100% (1)
- Azucar 1Document13 pagesAzucar 1Rafael PazNo ratings yet
- Epitelio 1Document33 pagesEpitelio 1Rafael PazNo ratings yet
- Tubulo 1Document5 pagesTubulo 1Rafael PazNo ratings yet
- Nucleo 1Document22 pagesNucleo 1Rafael PazNo ratings yet
- Sintesis 1Document27 pagesSintesis 1Rafael PazNo ratings yet
- Proteina 1Document17 pagesProteina 1Rafael PazNo ratings yet
- Enzima 1Document21 pagesEnzima 1Rafael PazNo ratings yet
- (FUERZA, ROBERT MORTON, BUENO) Training For Strength and Hypertrophy An Evidence-Based Approach ESDocument12 pages(FUERZA, ROBERT MORTON, BUENO) Training For Strength and Hypertrophy An Evidence-Based Approach ESJony CostaNo ratings yet
- Batagelj NMDocument79 pagesBatagelj NMJohanna Nieto HermosillaNo ratings yet
- A.I. Kapandji - #3 (Tronco y Raquis)Document46 pagesA.I. Kapandji - #3 (Tronco y Raquis)Fanii BellamyNo ratings yet
- Actividad de Nivelación 10 Segundo TrimestreDocument5 pagesActividad de Nivelación 10 Segundo TrimestreLUCY MALDONADO ROJASNo ratings yet
- Sistema OseoDocument5 pagesSistema OseoMaria Paula Martine DuqueNo ratings yet
- Medición de pliegues cutáneosDocument6 pagesMedición de pliegues cutáneosDeybi Torres0% (1)
- Concepto de Fuerza y Tipos de Fuerza en El Entrenamiento Muscular, DeportivoDocument17 pagesConcepto de Fuerza y Tipos de Fuerza en El Entrenamiento Muscular, DeportivoJuan Carlos GuevaraNo ratings yet
- Proyecto de Entrenamiento y Mejoramiento de La MemoriaDocument27 pagesProyecto de Entrenamiento y Mejoramiento de La MemoriaCecy Lezama100% (1)
- MuscularHistoDocument5 pagesMuscularHistoconche tumadreNo ratings yet
- Anteproyecto Del Filete de Cachama ApanadaDocument25 pagesAnteproyecto Del Filete de Cachama ApanadaCelia Coy PertuzNo ratings yet
- Guia de Musculación 2.0Document42 pagesGuia de Musculación 2.0jotarpzNo ratings yet
- BIOLOGÍA CuestionarioDocument5 pagesBIOLOGÍA CuestionarioJaniris EchavarriaNo ratings yet
- Funciones de Las Proteinas CuadroDocument2 pagesFunciones de Las Proteinas Cuadroverdeoros uaemexNo ratings yet
- Sistema de Comunicación de Los AnfibiosDocument8 pagesSistema de Comunicación de Los AnfibiosAlicia AlondracNo ratings yet
- Modulo de Estudio SubtenientesDocument47 pagesModulo de Estudio SubtenientesSTALINNo ratings yet
- Actividad 1 Plan de Prevención y Control de Patologías de Origen LaboralDocument21 pagesActividad 1 Plan de Prevención y Control de Patologías de Origen LaboralLuz Angela Henao Navarro100% (1)
- Pruebas Fisicas Policial NacionalDocument16 pagesPruebas Fisicas Policial NacionalAlex OwenNo ratings yet
- ACTIVIDAD 3 ESTUDIO DE CASO (1) EjemploDocument30 pagesACTIVIDAD 3 ESTUDIO DE CASO (1) EjemploMARIANo ratings yet
- TejidosVegetalesAnimalesDocument6 pagesTejidosVegetalesAnimalesVALERIE RODRÍGUEZNo ratings yet
- 9 Aparato LocomotorDocument35 pages9 Aparato Locomotormarcela gomezNo ratings yet
- Bases de Exploracion Neurologica Completa PDFDocument7 pagesBases de Exploracion Neurologica Completa PDFpiaxtlaNo ratings yet
- Efectos de la suplementación con creatina, ginseng y astrágalo sobre la fuerza y composición corporal en ancianosDocument11 pagesEfectos de la suplementación con creatina, ginseng y astrágalo sobre la fuerza y composición corporal en ancianosIsaias Acuña VerdugoNo ratings yet
- Cuerpo CiliarDocument13 pagesCuerpo CiliarRosario GarciaNo ratings yet
- BiomenicaDelCiclismo DiezSDocument13 pagesBiomenicaDelCiclismo DiezSnonograna75No ratings yet
- Enfermedades Del Sistema MuscularDocument5 pagesEnfermedades Del Sistema MuscularYelkys Aviles50% (2)