You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
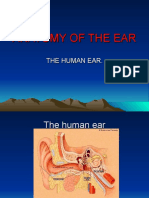
- 01.anatomy of The EarDocument52 pages01.anatomy of The EarJumanne Jay100% (1)
- Anatomy of Ear 1Document18 pagesAnatomy of Ear 1VikrantNo ratings yet
- Otoacoustic Emissions From MedscapeDocument5 pagesOtoacoustic Emissions From MedscapeMin-Joo Esther ParkNo ratings yet
- Dr. Karppasamy Notes PDFDocument275 pagesDr. Karppasamy Notes PDFRakshana ThangavelNo ratings yet
- CT Imaging of The Temporal Bone: An Anatomical Review With Illustrative Cases of CholesteatomasDocument116 pagesCT Imaging of The Temporal Bone: An Anatomical Review With Illustrative Cases of CholesteatomasDr-Firas Nayf Al-Thawabia100% (1)
- UnifloadDocument7 pagesUnifloadMacIqochgkjrtamlNo ratings yet
- DañlhsvDocument1 pageDañlhsvMacIqochgkjrtamlNo ratings yet
- 3Document4 pages3MacIqochgkjrtamlNo ratings yet
- Discos1: Grup/Intèrpret Títol EstilDocument28 pagesDiscos1: Grup/Intèrpret Títol EstilMacIqochgkjrtamlNo ratings yet
- Documento de Impresora Redirigido de Escritorio Remoto PDFDocument2 pagesDocumento de Impresora Redirigido de Escritorio Remoto PDFMacIqochgkjrtamlNo ratings yet
- AfjaobhauñfDocument9 pagesAfjaobhauñfMacIqochgkjrtamlNo ratings yet
- SaghDocument5 pagesSaghMacIqochgkjrtamlNo ratings yet
- AhsdkgjhañlrDocument17 pagesAhsdkgjhañlrMacIqochgkjrtamlNo ratings yet
- Plan Till A Flecha S CorazonDocument2 pagesPlan Till A Flecha S CorazonMacIqochgkjrtamlNo ratings yet
- Paw PatrolDocument8 pagesPaw PatrolMacIqochgkjrtamlNo ratings yet
- Otoacoustic Emission Suppression Testing A Clinician S Window Onto The Auditory Efferent Pathway PDFDocument12 pagesOtoacoustic Emission Suppression Testing A Clinician S Window Onto The Auditory Efferent Pathway PDFMacIqochgkjrtamlNo ratings yet
- Woodland Cake Tutorial: You Will NeedDocument13 pagesWoodland Cake Tutorial: You Will NeedMacIqochgkjrtaml100% (1)
- Plan Till A Flecha S CorazonDocument2 pagesPlan Till A Flecha S CorazonMacIqochgkjrtamlNo ratings yet
- DSI DYWIDAG ETA 13 0815 Post Tensioning System Using Strands enDocument62 pagesDSI DYWIDAG ETA 13 0815 Post Tensioning System Using Strands enMacIqochgkjrtamlNo ratings yet
- Plan Till A Flecha S CorazonDocument2 pagesPlan Till A Flecha S CorazonMacIqochgkjrtamlNo ratings yet
- Ergonomics 1 MVCDocument12 pagesErgonomics 1 MVCjamaica Malabuyoc100% (1)
- Anatomy and Physiology of Hearing SystemDocument63 pagesAnatomy and Physiology of Hearing SystemKharenza Vania Azarine Bachtiar100% (1)
- SensationDocument173 pagesSensationapi-261267976No ratings yet
- Project of Thesis - Listening ComprehensionDocument25 pagesProject of Thesis - Listening ComprehensionSonia Chauca50% (2)
- CongenitalDocument18 pagesCongenitalMatheus EmmanuelNo ratings yet
- Diseases of the EarDocument15 pagesDiseases of the Earsurg absiteNo ratings yet
- Hal 1-2Document4 pagesHal 1-2frita1986No ratings yet
- Ears 7. HearinglDocument11 pagesEars 7. HearingljottowagNo ratings yet
- Psy 100-Test 2Document59 pagesPsy 100-Test 2anon_964762231No ratings yet
- Special Senses Pathway: Visual, Auditory & VestibularDocument56 pagesSpecial Senses Pathway: Visual, Auditory & Vestibularmyka brilliant cristobalNo ratings yet
- OtosclerosisDocument85 pagesOtosclerosisigorNo ratings yet
- Tympanoplasty HandoutDocument2 pagesTympanoplasty HandoutNodsiriNo ratings yet
- SCIENCE: The Sense of Hearing What Are The Three Main Parts of The Human Ear?Document2 pagesSCIENCE: The Sense of Hearing What Are The Three Main Parts of The Human Ear?Fermina CachoNo ratings yet
- Hearing Anatomy OverviewDocument8 pagesHearing Anatomy OverviewMarie HwangNo ratings yet
- OtosclerosisDocument9 pagesOtosclerosisHelgaNo ratings yet
- Ear ExaminationDocument13 pagesEar Examinationsejal3vijNo ratings yet
- Discussion Paper On Hearing LossDocument36 pagesDiscussion Paper On Hearing LossAsha ShivaramNo ratings yet
- Asymmetrical Hearing Loss: Jessica PrasadDocument6 pagesAsymmetrical Hearing Loss: Jessica PrasadSujaya ChattopadhyayNo ratings yet
- Case Study - Special Senses and Reproductive SystemDocument2 pagesCase Study - Special Senses and Reproductive SystemGlydenne Glaire Poncardas GayamNo ratings yet
- Evaluation of The Patient With Vertigo - UpToDateDocument34 pagesEvaluation of The Patient With Vertigo - UpToDateadngdNo ratings yet
- Vestibular Evoked Myogenic Potentials: Preliminary ReportDocument8 pagesVestibular Evoked Myogenic Potentials: Preliminary ReportHerminaElenaNo ratings yet
- Management of Vertigo - DR - Attiya Rahma SpsDocument59 pagesManagement of Vertigo - DR - Attiya Rahma SpsHenni Pus Vera100% (1)
- Otosclerosis: Causes, Symptoms and TreatmentDocument25 pagesOtosclerosis: Causes, Symptoms and TreatmentHamid AliNo ratings yet
- Assessment of Deafness: Dr. Ajay Manickam JR - Dept of Ent RG Kar Medical CollegeDocument37 pagesAssessment of Deafness: Dr. Ajay Manickam JR - Dept of Ent RG Kar Medical CollegeDrravikumar BhandariNo ratings yet
- Vestibular NeuronitisDocument3 pagesVestibular NeuronitisAnish RajNo ratings yet