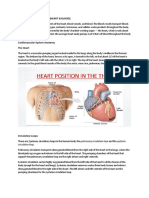
You might also like
- Nephrologi NotesDocument43 pagesNephrologi NotesSigit Harya HutamaNo ratings yet
- Lampiran Hasil SPSSSDocument3 pagesLampiran Hasil SPSSSSigit Harya HutamaNo ratings yet
- NIH Public Access: Inflammation and Wound Healing: The Role of The MacrophageDocument14 pagesNIH Public Access: Inflammation and Wound Healing: The Role of The MacrophageSigit Harya HutamaNo ratings yet
- Rat PDGF (Platelet-Derived Growth Factor) ELISA Kit PDFDocument9 pagesRat PDGF (Platelet-Derived Growth Factor) ELISA Kit PDFSigit Harya HutamaNo ratings yet
- Role of VEGF in Organogenesis: ReviewDocument10 pagesRole of VEGF in Organogenesis: ReviewSigit Harya HutamaNo ratings yet
- 144TD (C) 25 (E3) - Issue No 3 - Ear, Nose and Throat (ENT) Infections Antibiotic Guidelines PDFDocument9 pages144TD (C) 25 (E3) - Issue No 3 - Ear, Nose and Throat (ENT) Infections Antibiotic Guidelines PDFDellNo ratings yet
- Effects of Mesenchymal Stem Cells Conditioned Medium On Behavioral Aspects of Inflammatory Arthritic Pain Induced by Complete Freund's AdjuvantDocument9 pagesEffects of Mesenchymal Stem Cells Conditioned Medium On Behavioral Aspects of Inflammatory Arthritic Pain Induced by Complete Freund's AdjuvantSigit Harya HutamaNo ratings yet
- 7109 21855 1 SMDocument14 pages7109 21855 1 SMSigit Harya HutamaNo ratings yet
- Experimental and Toxicologic PathologyDocument6 pagesExperimental and Toxicologic PathologySigit Harya HutamaNo ratings yet
- Human Mesenchymal Stem Cell-Conditioned Medium Improves Cardiac Function Following Myocardial InfarctionDocument9 pagesHuman Mesenchymal Stem Cell-Conditioned Medium Improves Cardiac Function Following Myocardial InfarctionSigit Harya HutamaNo ratings yet
- Rat IL-10 (Interleukin-10) ELISA KitDocument9 pagesRat IL-10 (Interleukin-10) ELISA KitSigit Harya HutamaNo ratings yet
- Rat PDGF ELISA Kit Instruction ManualDocument9 pagesRat PDGF ELISA Kit Instruction ManualSigit Harya HutamaNo ratings yet
- Passing Grade SBMPTN 2016Document124 pagesPassing Grade SBMPTN 2016Aprilia Ulifatu FadilaNo ratings yet
- Amin 2015Document9 pagesAmin 2015Sigit Harya HutamaNo ratings yet
- 18Document1 page18Sigit Harya HutamaNo ratings yet
- Molecular Sciences: An Open Access JournalDocument2 pagesMolecular Sciences: An Open Access JournalSigit Harya HutamaNo ratings yet
- 14Document1 page14Sigit Harya HutamaNo ratings yet
- Jurnal Dermatitis Hong, 2014Document2 pagesJurnal Dermatitis Hong, 2014Sigit Harya HutamaNo ratings yet
- Mesenchymal Stem Cell Therapy in Type Diabetes Mellitus and Its Main Complications From Experimental Findings To Clinical Practice 2157 7633.1000227Document10 pagesMesenchymal Stem Cell Therapy in Type Diabetes Mellitus and Its Main Complications From Experimental Findings To Clinical Practice 2157 7633.1000227Sigit Harya HutamaNo ratings yet
- Ijeb Inst Auth PDFDocument2 pagesIjeb Inst Auth PDFSigit Harya HutamaNo ratings yet
- Relationship Between Environmental Sanitation and Malaria IncidenceDocument7 pagesRelationship Between Environmental Sanitation and Malaria IncidenceSigit Harya HutamaNo ratings yet
- znl1000 PDFDocument6 pagesznl1000 PDFSigit Harya HutamaNo ratings yet
- Nihms266207 PDFDocument19 pagesNihms266207 PDFSigit Harya HutamaNo ratings yet
- 3Document16 pages3Sigit Harya HutamaNo ratings yet
- 2227Document9 pages2227Faizah Min FadhlillahNo ratings yet
- Delegates Proposal + Invitation LetterDocument38 pagesDelegates Proposal + Invitation LetterSigit Harya HutamaNo ratings yet
- 2227Document9 pages2227Faizah Min FadhlillahNo ratings yet
- Local Production of Secretory Iga in The Eye-Associated Lymphoid Tissue (Ealt) of The Normal Human Ocular SurfaceDocument8 pagesLocal Production of Secretory Iga in The Eye-Associated Lymphoid Tissue (Ealt) of The Normal Human Ocular SurfaceSigit Harya HutamaNo ratings yet
- Unexpected Role of Inflammatory Signaling In.5Document6 pagesUnexpected Role of Inflammatory Signaling In.5Sigit Harya HutamaNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5783)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (72)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Cardiovascular SystemDocument17 pagesCardiovascular SystemPoojaNo ratings yet
- The SupernaturalistDocument2 pagesThe SupernaturalistAlex LuNo ratings yet
- 6 135Document6 pages6 135Ashok LenkaNo ratings yet
- Plants and EnvironmentDocument284 pagesPlants and EnvironmentYen LeNo ratings yet
- Fertilization to Birth: A Guide to Pregnancy DevelopmentDocument7 pagesFertilization to Birth: A Guide to Pregnancy DevelopmentSamson BongsiwNo ratings yet
- Tissue processing problems and artifactsDocument18 pagesTissue processing problems and artifactsOsama Bakheet100% (2)
- 20-Advances in Cognitive TheoryDocument24 pages20-Advances in Cognitive TheoryAndrea ZeballosNo ratings yet
- The Thromboelastogram (TEG®) : A Five-Minute Primer For The Emergency PhysicianDocument11 pagesThe Thromboelastogram (TEG®) : A Five-Minute Primer For The Emergency PhysicianMarcelliaNo ratings yet
- Anti AIDS HerbsDocument6 pagesAnti AIDS Herbskhansa maryamNo ratings yet
- Nivell C1 Anglés / Nivel C1 Inglés Comprensió Oral / Comprensión OralDocument8 pagesNivell C1 Anglés / Nivel C1 Inglés Comprensió Oral / Comprensión OralHiNo ratings yet
- Algal Biorefinery - An Integrated Approach For Sustainable Biodiesel ProductionDocument16 pagesAlgal Biorefinery - An Integrated Approach For Sustainable Biodiesel ProductionCarlos GamarraNo ratings yet
- Adenium PropogationDocument1 pageAdenium PropogationTanNo ratings yet
- FleaDocument23 pagesFleaAAREEZ IMRAANNo ratings yet
- A2 Biology Notes For EdexcelDocument150 pagesA2 Biology Notes For EdexcelAbdulrahman Jijawi83% (6)
- 20 Amino Acids in Human ProteinDocument2 pages20 Amino Acids in Human Proteinpanomo nasabyNo ratings yet
- Application Summary Sheet For Programme-Specific Requirements and Selection: Master's Programme in BiopharmaceuticalsDocument4 pagesApplication Summary Sheet For Programme-Specific Requirements and Selection: Master's Programme in BiopharmaceuticalsZikoraNo ratings yet
- Cambridge International AS and A Level Biology Revision Guide PDFDocument40 pagesCambridge International AS and A Level Biology Revision Guide PDFGirish Krishnan67% (3)
- Philippine Threatened Fauna Booklet Volume 1 PDFDocument120 pagesPhilippine Threatened Fauna Booklet Volume 1 PDFAloy DuyaNo ratings yet
- Chandini ThesisDocument71 pagesChandini Thesismallesh D trigarNo ratings yet
- Micro Biology EMDocument320 pagesMicro Biology EMdeliciaammuNo ratings yet
- DissertationDocument7 pagesDissertationChandra SekharNo ratings yet
- Protocolo de Extracción de ADN Bacteriano - AusubelDocument5 pagesProtocolo de Extracción de ADN Bacteriano - AusubelLesly CastilloNo ratings yet
- Formulir Review Judul Skripsi 2019 (Update 28 Maret 2019) Kevin ThenediDocument2 pagesFormulir Review Judul Skripsi 2019 (Update 28 Maret 2019) Kevin ThenediKevin ThenediNo ratings yet
- 2019PSNLLB43642-648Syahdietal 1 PDFDocument15 pages2019PSNLLB43642-648Syahdietal 1 PDFAlina Vitali TabarceaNo ratings yet
- Bmich 2Document1 pageBmich 2umahsuthanNo ratings yet
- Mothership Character SheetDocument2 pagesMothership Character SheetAlexander Dalton0% (1)
- Cellular Memory and How It WorksDocument8 pagesCellular Memory and How It Worksbiba 4life100% (1)
- Physical Basics of Informational InteractionDocument45 pagesPhysical Basics of Informational Interactiondb1970100% (1)
- Cystic Fibrosis: Nature Reviews Disease Primers May 2015Document20 pagesCystic Fibrosis: Nature Reviews Disease Primers May 2015Tapan AnkleshwariaNo ratings yet
- Physiology Gastric EmptyingDocument14 pagesPhysiology Gastric EmptyingyorleNo ratings yet