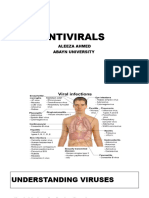
You might also like
- Renny Soal Pas 9Document1 pageRenny Soal Pas 9Yasinta Swastika AyuNo ratings yet
- DAPUSDocument1 pageDAPUSYasinta Swastika AyuNo ratings yet
- How to set up water dispenserDocument3 pagesHow to set up water dispenserYasinta Swastika AyuNo ratings yet
- How to set up water dispenserDocument3 pagesHow to set up water dispenserYasinta Swastika AyuNo ratings yet
- Penyakit Inf Saluran Pencernaan PDFDocument9 pagesPenyakit Inf Saluran Pencernaan PDFAde PermanaNo ratings yet
- Hasil Pengamatan SintaDocument8 pagesHasil Pengamatan SintaYasinta Swastika AyuNo ratings yet
- Its A Wrap!Document1 pageIts A Wrap!Yasinta Swastika AyuNo ratings yet
- ANJENGDocument1 pageANJENGYasinta Swastika AyuNo ratings yet
- Abstract (1) SorgumDocument2 pagesAbstract (1) SorgumYasinta Swastika AyuNo ratings yet
- TranspirationDocument10 pagesTranspirationYasinta Swastika AyuNo ratings yet
- Ujian Praktek Bahasa Inggris Kelas 9 TH Ajaran 2011/2012 SMP Negeri 4 MalangDocument1 pageUjian Praktek Bahasa Inggris Kelas 9 TH Ajaran 2011/2012 SMP Negeri 4 MalangYasinta Swastika AyuNo ratings yet
- Zaitsev, Y.2001.Exotic Zaitsev, Y.2001.Exotic Zaitsev, Y.2001.Exotic Zaitsev, Y.2001.ExoticDocument4 pagesZaitsev, Y.2001.Exotic Zaitsev, Y.2001.Exotic Zaitsev, Y.2001.Exotic Zaitsev, Y.2001.ExoticYasinta Swastika AyuNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (890)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- NERO215 Handout 3Document4 pagesNERO215 Handout 3Heena GoyalNo ratings yet
- Jamb Syllabus For BiologyDocument26 pagesJamb Syllabus For BiologyFaithNo ratings yet
- Introduction To Biological Evolution (Table of Contents)Document10 pagesIntroduction To Biological Evolution (Table of Contents)Mitch Forbes100% (1)
- Lec 10 AntiviralsDocument26 pagesLec 10 AntiviralsAiqa QaziNo ratings yet
- Intro To Taxonomy StudentDocument30 pagesIntro To Taxonomy StudentAdeyemi AdamsNo ratings yet
- CARBOHYDRATE ABSORPTION (Ref)Document22 pagesCARBOHYDRATE ABSORPTION (Ref)NEERAJA S RAJNo ratings yet
- Effects of Androgen on Male and Female ChicksDocument22 pagesEffects of Androgen on Male and Female ChicksMarie Elexis RevillezaNo ratings yet
- Hyperthermia SystemicDocument21 pagesHyperthermia Systemicataraix22No ratings yet
- Genes Hearing DeafnessDocument328 pagesGenes Hearing DeafnessjvojnicNo ratings yet
- Collagen in Wound Healing: BioengineeringDocument15 pagesCollagen in Wound Healing: BioengineeringMarco Antonio Garcia GonzalezNo ratings yet
- VirologyLearnTable 2017-Sketchy PDFDocument4 pagesVirologyLearnTable 2017-Sketchy PDFdfgbdfgNo ratings yet
- Oxidative Stress-Induced Hair Growth InhibitionDocument10 pagesOxidative Stress-Induced Hair Growth InhibitionDavid Fernando Rojas BayonaNo ratings yet
- Uncovering Principles of Brain EvolutionDocument36 pagesUncovering Principles of Brain EvolutionCulture HeritageNo ratings yet
- New Insights in Dialysis Membrane BiocompatibilityDocument7 pagesNew Insights in Dialysis Membrane BiocompatibilityDani ursNo ratings yet
- Cluster IIDocument11 pagesCluster IITJ OberioNo ratings yet
- GCUF Final Exams Date SheetDocument20 pagesGCUF Final Exams Date SheetAwais ChaudharyNo ratings yet
- Cabison - Histopath ReviewerDocument17 pagesCabison - Histopath ReviewerPatricia CabisonNo ratings yet
- Threadfins of The World (Family Polynemidae)Document8 pagesThreadfins of The World (Family Polynemidae)cavrisNo ratings yet
- Experiment No. 2 Amino Acids and Proteins (Part I)Document12 pagesExperiment No. 2 Amino Acids and Proteins (Part I)Anne GellieNo ratings yet
- Essay GENETICALLY MODIFIED FOODDocument7 pagesEssay GENETICALLY MODIFIED FOODSandro BomfimNo ratings yet
- Chapter 9 - Plant Biology: MarkschemeDocument17 pagesChapter 9 - Plant Biology: MarkschemeAtulya BharadwajNo ratings yet
- Bacterial GrowthDocument16 pagesBacterial GrowthSaudah IbrahimNo ratings yet
- Photosynthesis PDFDocument30 pagesPhotosynthesis PDFhumble happyNo ratings yet
- Chapter 22Document91 pagesChapter 22YuoMatsukiNo ratings yet
- Histology SlidesDocument26 pagesHistology SlidesDhruvJaiswal100% (1)
- N20 H2 P2 Answers-1Document10 pagesN20 H2 P2 Answers-1Samuel TeohNo ratings yet
- OkuobakaDocument29 pagesOkuobakaEricsson BotchweyNo ratings yet
- Act 2. Physiology Lab. (Yeo's Part)Document3 pagesAct 2. Physiology Lab. (Yeo's Part)YEO, REGGIE ALBERT A.No ratings yet
- Engineering 3d Tissue Test SystemsDocument435 pagesEngineering 3d Tissue Test SystemsDANIEL FELIPE RINCON OVALLENo ratings yet
- Propagación de Plantas Por SemillaDocument50 pagesPropagación de Plantas Por Semillamaquita1No ratings yet