You might also like
- TerapiaDocument5 pagesTerapiaCentri CopiasNo ratings yet
- Masaje para Cicatriz..Document16 pagesMasaje para Cicatriz..anon_287230938100% (1)
- Ergonomia Ambiental PresentacionDocument46 pagesErgonomia Ambiental PresentacionGenesis J. Vallejos100% (1)
- Monografia de Colesterol y TrigliceridosDocument23 pagesMonografia de Colesterol y TrigliceridosLiz M VillaVqz100% (1)
- Segundos MensajerosDocument42 pagesSegundos MensajerosMitch Peraza80% (10)
- Historia ClínicaDocument4 pagesHistoria ClínicaOscar GomezNo ratings yet
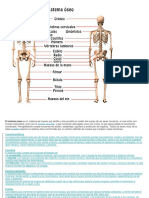
- Sistema OseoDocument13 pagesSistema OseoLeila OrtizNo ratings yet
- El Científico Robert Brawn Fue Quien Descubrió El Núcleo de La Célula en El Año 1665. Nota CientíficaDocument3 pagesEl Científico Robert Brawn Fue Quien Descubrió El Núcleo de La Célula en El Año 1665. Nota CientíficaIvan Tacza SantanaNo ratings yet
- Determinación ColesterolDocument13 pagesDeterminación ColesterolNicolas BustamanteNo ratings yet
- Reflejos ArcaicosDocument35 pagesReflejos ArcaicosDani Estrada100% (1)
- CalidadVidaCáncer40Document3 pagesCalidadVidaCáncer40Alejandra Luna100% (3)
- El Ion Calcio - Su Regulación y Función en La Célula SS PancreáticaDocument19 pagesEl Ion Calcio - Su Regulación y Función en La Célula SS PancreáticaHarold Rodriguez EcheverryNo ratings yet
- La Transmicion NerviosaDocument7 pagesLa Transmicion Nerviosaeduardo100% (2)
- R ISADocument5 pagesR ISAluz castilloNo ratings yet
- El Mecanismo de Acoplamiento ExcitaciónDocument4 pagesEl Mecanismo de Acoplamiento ExcitaciónJuan AguilarNo ratings yet
- Daryenel Huitron Autocoides y Segundos MensajerosDocument22 pagesDaryenel Huitron Autocoides y Segundos MensajerosDaryenel HuitrónNo ratings yet
- Acoplamineto Excitación - ContracciónDocument4 pagesAcoplamineto Excitación - ContracciónKim Hyun JoongNo ratings yet
- Función Del Calcio Como Mensajero IntracelularDocument14 pagesFunción Del Calcio Como Mensajero IntracelularRegina Lozada ChaconNo ratings yet
- Calcio gluconato: electrolito esencialDocument13 pagesCalcio gluconato: electrolito esencialCynthia Quispe CiezaNo ratings yet
- Antagonistas Del Calcio en La Hipertensión ArterialDocument22 pagesAntagonistas Del Calcio en La Hipertensión ArterialedisonNo ratings yet
- Farmacodinamia del músculo liso visceral: mecanismos de contracción y relajaciónDocument22 pagesFarmacodinamia del músculo liso visceral: mecanismos de contracción y relajaciónCarina VillegaNo ratings yet
- Preguntes FISIODocument8 pagesPreguntes FISIOmargotNo ratings yet
- Canales IónicosDocument22 pagesCanales IónicosMika SuspiroNo ratings yet
- Control de La Función CardiovascularDocument10 pagesControl de La Función CardiovascularMartin Ojeda CarpioNo ratings yet
- Potencial de Accion CardiacoDocument8 pagesPotencial de Accion CardiacoAngelo Anaya CaponeNo ratings yet
- Miastenia Gravis (Referencias y Preguntas)Document7 pagesMiastenia Gravis (Referencias y Preguntas)plopezh1No ratings yet
- Taller SeñalizaciónDocument5 pagesTaller Señalizaciónrichard urreaNo ratings yet
- La ATPasa de calcio en el sistema nerviosoDocument8 pagesLa ATPasa de calcio en el sistema nerviosoeliocesar92No ratings yet
- Trastornos Del Equilibrio ElectroliticoDocument63 pagesTrastornos Del Equilibrio ElectroliticoFernanda Pineda GeaNo ratings yet
- IjojioiuiuDocument8 pagesIjojioiuiuArlyNo ratings yet
- Fisiología membrana celular y potenciales de acciónDocument29 pagesFisiología membrana celular y potenciales de acciónElizabethNo ratings yet
- Eaxmen de FisioDocument5 pagesEaxmen de FisioWilmar YPeNo ratings yet
- Fisiología de La Contracción CardiacaDocument30 pagesFisiología de La Contracción CardiacaHebert Dario RodriguezNo ratings yet
- Páncreas EndócrinoDocument8 pagesPáncreas Endócrinoyulianaledesmantvg16No ratings yet
- Comunicación celular en el SNCDocument2 pagesComunicación celular en el SNCwilson albornozNo ratings yet
- Corrientes IonicasDocument6 pagesCorrientes Ionicasoscar olivasNo ratings yet
- Canales de CaDocument14 pagesCanales de CaMiguel MendozaNo ratings yet
- Acomplamiento Excito-ContractilDocument4 pagesAcomplamiento Excito-ContractilFedericoNo ratings yet
- Guía de Estudio s3 Bio DiciembreDocument7 pagesGuía de Estudio s3 Bio DiciembreJosefina EchandiNo ratings yet
- Ruiz - Parte Del Inf. Memb. basolaterALDocument9 pagesRuiz - Parte Del Inf. Memb. basolaterALJean VillanuevaNo ratings yet
- AcetilcolinaDocument9 pagesAcetilcolinakatya monserrat cocolepsi fortizNo ratings yet
- TEMA 4 FISIODocument6 pagesTEMA 4 FISIOAngel ChavesNo ratings yet
- Caso 12 - BCM - Una Situación DesequilibranteDocument5 pagesCaso 12 - BCM - Una Situación DesequilibrantecmlgarinNo ratings yet
- Preguntas Del Pase de VisitaDocument7 pagesPreguntas Del Pase de VisitaIvet SuarezNo ratings yet
- EstructuraDocument4 pagesEstructurajesus perezNo ratings yet
- Bomba Sodio-PotacioDocument7 pagesBomba Sodio-PotacioJose Anthony GarciaNo ratings yet
- Farmacodinamia 1Document12 pagesFarmacodinamia 1Nahuel RodriguezNo ratings yet
- Transducción de SeñalesDocument31 pagesTransducción de SeñalesBelén JiménezNo ratings yet
- Fisiología de Los Tejidos ExcitablesDocument52 pagesFisiología de Los Tejidos ExcitablesJosé CarlosNo ratings yet
- Banco de Fisio ActualizadoDocument11 pagesBanco de Fisio ActualizadoNikolas SeasNo ratings yet
- CalcioDocument3 pagesCalcioflorenciarenieroNo ratings yet
- Sildenafil - Harper BioquimicaDocument4 pagesSildenafil - Harper BioquimicaF FretesNo ratings yet
- Actividad SinápticaDocument48 pagesActividad SinápticaGadiel Couoh RosadoNo ratings yet
- Seminario EndoDocument6 pagesSeminario EndogdomingufNo ratings yet
- Respuestas Celulares Post ReceptorDocument12 pagesRespuestas Celulares Post ReceptorBrandon Stewart EspinozaNo ratings yet
- Trastornos Del Potasio Nefrología Al DíaDocument1 pageTrastornos Del Potasio Nefrología Al DíaXavier TorresNo ratings yet
- Receptores de Glutamato y Renina AngiotensinaDocument19 pagesReceptores de Glutamato y Renina AngiotensinaYunuen Ivonne Martínez RodríguezNo ratings yet
- Receptor Guanililciclasa. GMPc.Document32 pagesReceptor Guanililciclasa. GMPc.abanto90No ratings yet
- Enfermedades asociadas con mutaciones SERCADocument9 pagesEnfermedades asociadas con mutaciones SERCALuisaGonzalezNo ratings yet
- Segundos MensajerosDocument14 pagesSegundos MensajerosArnold Gomez AlvaNo ratings yet
- NeurotransmisoresDocument72 pagesNeurotransmisoresAyuwoki Hee heeNo ratings yet
- Tp2 Garcia RosaDocument8 pagesTp2 Garcia Rosarosa maria garciaNo ratings yet
- Asignación SINAPSIS y NEUROTRANSMISORESDocument7 pagesAsignación SINAPSIS y NEUROTRANSMISORESPaola AlmonteNo ratings yet
- Cuaderno FarmacoDocument14 pagesCuaderno Farmacoalejandro acosta tovarNo ratings yet
- Electrofisiología Del Miocito VentricularDocument4 pagesElectrofisiología Del Miocito VentricularUSMP FN ARCHIVOSNo ratings yet
- Desórdenes en Receptores CelularesDocument40 pagesDesórdenes en Receptores CelularesMarco MadridNo ratings yet
- Clase 7 Canales y Transducción CelularDocument37 pagesClase 7 Canales y Transducción CelularCristina FavelaNo ratings yet
- Canales Ionicos Algo K Avance PZZZ Kelo MemeDocument25 pagesCanales Ionicos Algo K Avance PZZZ Kelo MemeMiluska Liza NiquenNo ratings yet
- Transmision SinapticaDocument8 pagesTransmision SinapticaALICIA LISETH VENTURA TINEONo ratings yet
- Hipotesis AntibioticosDocument9 pagesHipotesis AntibioticosLiz M VillaVqzNo ratings yet
- Expo de CalidadDocument12 pagesExpo de CalidadLiz M VillaVqzNo ratings yet
- fARMACOBOTANICA INFormeDocument13 pagesfARMACOBOTANICA INFormeLiz M VillaVqzNo ratings yet
- Factores Que ModificanDocument23 pagesFactores Que ModificanLiz M VillaVqzNo ratings yet
- Gestión Del Talento HumanoDocument2 pagesGestión Del Talento HumanoLiz M VillaVqzNo ratings yet
- Etica y Responsabilidad SocialDocument10 pagesEtica y Responsabilidad SocialLiz M VillaVqzNo ratings yet
- FORTALEZASDocument2 pagesFORTALEZASLiz M VillaVqzNo ratings yet
- Clinica .GeneralidadesDocument9 pagesClinica .GeneralidadesLiz M VillaVqzNo ratings yet
- Farmacoquimica I PracticaDocument2 pagesFarmacoquimica I PracticaLiz M VillaVqzNo ratings yet
- Practica N 4 CasiDocument4 pagesPractica N 4 CasiLiz M VillaVqzNo ratings yet
- Clinica .GeneralidadesDocument9 pagesClinica .GeneralidadesLiz M VillaVqzNo ratings yet
- Análisis Clínico II HormonasDocument7 pagesAnálisis Clínico II HormonasLiz M VillaVqzNo ratings yet
- Grupo DeontologiaDocument2 pagesGrupo DeontologiaLiz M VillaVqzNo ratings yet
- Ácido AcetilsalicílicoDocument1 pageÁcido AcetilsalicílicoLiz M VillaVqzNo ratings yet
- MecanismpDocument2 pagesMecanismpLiz M VillaVqzNo ratings yet
- Cuadro ResumenDocument5 pagesCuadro ResumenLiz M VillaVqzNo ratings yet
- HTTPDocument1 pageHTTPLiz M VillaVqzNo ratings yet
- Informe #1 ClinicoDocument17 pagesInforme #1 ClinicoLiz M VillaVqzNo ratings yet
- LEVOSULPIRIDADocument1 pageLEVOSULPIRIDALiz M VillaVqzNo ratings yet
- Informe #3 Analisis Clinico IIDocument13 pagesInforme #3 Analisis Clinico IILiz M VillaVqz50% (2)
- Informe #2 Analisis Clinico IIDocument15 pagesInforme #2 Analisis Clinico IILiz M VillaVqzNo ratings yet
- Tema de AnalisisDocument1 pageTema de AnalisisLiz M VillaVqzNo ratings yet
- Informe #9 de BioquimicaDocument16 pagesInforme #9 de BioquimicaLiz M VillaVqzNo ratings yet
- Cuadro ResumenDocument5 pagesCuadro ResumenLiz M VillaVqzNo ratings yet
- Informe #9 de BioquimicaDocument16 pagesInforme #9 de BioquimicaLiz M VillaVqzNo ratings yet
- Arandano ANIDocument24 pagesArandano ANILiz M VillaVqzNo ratings yet
- Determinacion de Grupo RHDocument1 pageDeterminacion de Grupo RHLiz M VillaVqzNo ratings yet
- Nuevo InformeDocument1 pageNuevo InformeLiz M VillaVqzNo ratings yet
- Medicina Legal FILIACIONDocument3 pagesMedicina Legal FILIACIONAlexandra J Delgado GNo ratings yet
- Formacion Civica y Etica 5Document5 pagesFormacion Civica y Etica 5Minerva PerezNo ratings yet
- Monografia de Control de CalidadDocument43 pagesMonografia de Control de CalidadAlejandro MenesesNo ratings yet
- Correlaciones Clínicas Del Hipotálamo, Hipófisis, y Amígdala PDFDocument25 pagesCorrelaciones Clínicas Del Hipotálamo, Hipófisis, y Amígdala PDFCésar Augusto Gonzales QuiñonesNo ratings yet
- Asma infantil: guía actualizada de diagnóstico y tratamientoDocument38 pagesAsma infantil: guía actualizada de diagnóstico y tratamientoRenzo D Al FaNo ratings yet
- Genes LCT y Mcm6. - Ivami - AcDocument2 pagesGenes LCT y Mcm6. - Ivami - Acalvaro acNo ratings yet
- Virología T2 Apuntes 09-02Document7 pagesVirología T2 Apuntes 09-02kittycatcegoNo ratings yet
- Cuadernillo Defectos Geneticos 2017Document36 pagesCuadernillo Defectos Geneticos 2017Raizen GrengerNo ratings yet
- Hemograma CompletoDocument39 pagesHemograma CompletoWilliam Apaza MamaniNo ratings yet
- PROTEÍNASDocument29 pagesPROTEÍNASLuz Elena Collado ArapaNo ratings yet
- Diapositiva de DaltonismoDocument9 pagesDiapositiva de DaltonismoEdgar Godoy100% (2)
- Institución Educativa Distrital Salvador EntregasDocument17 pagesInstitución Educativa Distrital Salvador EntregasJesus HenríquezNo ratings yet
- Sistemas Sensoriales (Visión, Audición, Olfato, Gusto y Tacto) - Sesion Nro 09Document13 pagesSistemas Sensoriales (Visión, Audición, Olfato, Gusto y Tacto) - Sesion Nro 09JorgeVallejoCamachoNo ratings yet
- Pedic Ulos IsDocument18 pagesPedic Ulos IsRosario Medalit Quezada PacoraNo ratings yet
- Esguince CervicalDocument6 pagesEsguince CervicalFercho YañezNo ratings yet
- Costo de Digestión Del Murciélago Pescador Myotis VivesiDocument21 pagesCosto de Digestión Del Murciélago Pescador Myotis VivesiNataly Priscila MendezNo ratings yet
- Energia en MonogastricosDocument5 pagesEnergia en MonogastricosAngie ParedesNo ratings yet
- Biología de la creencia LiptonDocument2 pagesBiología de la creencia LiptonKvn CardenasNo ratings yet
- Alteraciones de la pigmentación cutánea: Melasma y vitiligoDocument43 pagesAlteraciones de la pigmentación cutánea: Melasma y vitiligoNaty LastraNo ratings yet
- Vitamina KDocument18 pagesVitamina Kdeisy_jasmin17No ratings yet
- Embarazo Multiple 10 HojasDocument11 pagesEmbarazo Multiple 10 HojasPercy Reyes MerinoNo ratings yet
- PKU: FENILCETONURIADocument19 pagesPKU: FENILCETONURIAKevin David Galindo CardosoNo ratings yet