You might also like
- 2009 - Dna Fragm and Pregnancy LossDocument8 pages2009 - Dna Fragm and Pregnancy LossMaría ElenaNo ratings yet
- Eren Preiss 2006Document19 pagesEren Preiss 2006María ElenaNo ratings yet
- 2009 - Macs Clinical OutcomeDocument7 pages2009 - Macs Clinical OutcomeMaría ElenaNo ratings yet
- 2002 - Role of Sperm Function in Failed FertilizationDocument4 pages2002 - Role of Sperm Function in Failed FertilizationMaría ElenaNo ratings yet
- 10 1080@01485010050193995Document11 pages10 1080@01485010050193995María ElenaNo ratings yet
- Does Human Sperm Nuclear DNA Integrity Affect Embryo Quality?Document7 pagesDoes Human Sperm Nuclear DNA Integrity Affect Embryo Quality?María ElenaNo ratings yet
- Peart2010 PDFDocument2 pagesPeart2010 PDFMaría ElenaNo ratings yet
- 2002 - Role of Sperm Function in Failed FertilizationDocument6 pages2002 - Role of Sperm Function in Failed FertilizationMaría ElenaNo ratings yet
- Johnson2010 PDFDocument16 pagesJohnson2010 PDFMaría ElenaNo ratings yet
- Function of Sperm Chromatin Structural Elements in Fertilization and DevelopmentDocument7 pagesFunction of Sperm Chromatin Structural Elements in Fertilization and DevelopmentMaría ElenaNo ratings yet
- Minireview Deoxyribonucleic Acid Loop Domain Tertiary Structure in Mammalian Spermatozoa'Document9 pagesMinireview Deoxyribonucleic Acid Loop Domain Tertiary Structure in Mammalian Spermatozoa'María ElenaNo ratings yet
- E Vector PatternDocument5 pagesE Vector PatternMaría ElenaNo ratings yet
- A Model For The Function of Sperm DNA Degradation: Monika A. Ward and W. Steven WardDocument8 pagesA Model For The Function of Sperm DNA Degradation: Monika A. Ward and W. Steven WardMaría ElenaNo ratings yet
- Dynamics of The Sperm NucleusDocument7 pagesDynamics of The Sperm NucleusMaría ElenaNo ratings yet
- 2002 - Role of Sperm Function in Failed FertilizationDocument6 pages2002 - Role of Sperm Function in Failed FertilizationMaría ElenaNo ratings yet
- SpermogramDocument286 pagesSpermogramalmutazimNo ratings yet
- Bees, Daphnia, and Polarized LightDocument5 pagesBees, Daphnia, and Polarized LightMaría ElenaNo ratings yet
- Honeybee Naviagtion. Following Routes Using Polarized-Light CuesDocument6 pagesHoneybee Naviagtion. Following Routes Using Polarized-Light CuesMaría ElenaNo ratings yet
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (894)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (265)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- DL-MPS901A Flexible Manufacturing System Training GuideDocument118 pagesDL-MPS901A Flexible Manufacturing System Training Guidethanhga22No ratings yet
- Understanding Quantum Physics Foundations BTech First Year Engineering Physics LectureDocument45 pagesUnderstanding Quantum Physics Foundations BTech First Year Engineering Physics LectureChop DownNo ratings yet
- A Detailed Lesson Plan in Arts IV - Color Harmony: Hipolito, MariellaDocument22 pagesA Detailed Lesson Plan in Arts IV - Color Harmony: Hipolito, MariellaMariella Monique100% (1)
- EAPP PeriodicalDocument8 pagesEAPP PeriodicalDherick RaleighNo ratings yet
- SEC - 04 - Security LightingDocument20 pagesSEC - 04 - Security LightingAnonymous bKp9kE0DYNo ratings yet
- (K14) - Contact LensesDocument33 pages(K14) - Contact LensesSyarifah FauziahNo ratings yet
- Case ReportDocument26 pagesCase ReportDesti EryaniNo ratings yet
- New Research Papers in PhysicsDocument8 pagesNew Research Papers in Physicsafeawjjwp100% (1)
- Alternate Light Source Imaging Forensic Photography TechniquesDocument99 pagesAlternate Light Source Imaging Forensic Photography Techniquesjadrio1556No ratings yet
- Ex-Proof Floodlight - Cutsheet (NJZ-FEL-B)Document7 pagesEx-Proof Floodlight - Cutsheet (NJZ-FEL-B)anderNo ratings yet
- Short Stories British Tales Isaac Newton AnswersDocument2 pagesShort Stories British Tales Isaac Newton AnswersEva Rico100% (1)
- Graduation Project PV English VersionDocument56 pagesGraduation Project PV English VersionabidineNo ratings yet
- Scott Westerfeld - Midnighters 03 - Blue NoonDocument321 pagesScott Westerfeld - Midnighters 03 - Blue NoonIvan MartNo ratings yet
- Intelligent Solar Street Light System Using SensorsDocument4 pagesIntelligent Solar Street Light System Using SensorsmnotcoolNo ratings yet
- WLB32 Industrial LED Light Bar (AC) : DatasheetDocument8 pagesWLB32 Industrial LED Light Bar (AC) : DatasheetDanono PadillaNo ratings yet
- Gots Approved Chemicals List 2015Document1,035 pagesGots Approved Chemicals List 2015Hrishikesh Dhawadshikar56% (9)
- Paulo Correa and Alexandra Correa - Experimental Aetherometry Volume IIA IntroductionDocument9 pagesPaulo Correa and Alexandra Correa - Experimental Aetherometry Volume IIA IntroductionSonyRed100% (1)
- Ocular Health History SheetDocument1 pageOcular Health History SheetSafa Abdualrahaman Ali HamadNo ratings yet
- Conclusions & InferencesDocument69 pagesConclusions & InferencesPaulina ReyesNo ratings yet
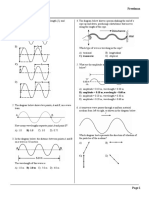
- Waves Probs 99-05 Answer KeyDocument22 pagesWaves Probs 99-05 Answer KeyRica RepdosNo ratings yet
- Kabul Image April 2011Document27 pagesKabul Image April 2011Kabul ImageNo ratings yet
- 25 Color Coding TelephoneDocument2 pages25 Color Coding TelephoneEkimYacamNo ratings yet
- Led 0606Document32 pagesLed 0606Alex Lim100% (1)
- Thermal-Electromagnetic Coupled System Simulator Using Transmission-Line Matrix (TLM)Document9 pagesThermal-Electromagnetic Coupled System Simulator Using Transmission-Line Matrix (TLM)TI Journals PublishingNo ratings yet
- The Electromagnetic SpectrumDocument25 pagesThe Electromagnetic SpectrumMark Niño JavierNo ratings yet
- FlexScan 1807 Gen BroDocument13 pagesFlexScan 1807 Gen BroSpicNo ratings yet
- DAY 5 HTML Quotation and Citation ElementsDocument38 pagesDAY 5 HTML Quotation and Citation ElementsNeil Christian CeballosNo ratings yet
- October November 23 Paper 12Document16 pagesOctober November 23 Paper 12avanishh914No ratings yet
- Optician Training Manual 4thDocument280 pagesOptician Training Manual 4thSuresh Nair100% (6)