You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Human Brain: An Introduction To Its Functional Anatomy - John NolteDocument733 pagesThe Human Brain: An Introduction To Its Functional Anatomy - John NolteNicolasDarmanthé100% (5)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- No Longer Living The Sweet Life: Losing Control of Diabetes: Part I - Annual ConsultationDocument9 pagesNo Longer Living The Sweet Life: Losing Control of Diabetes: Part I - Annual Consultationpedro buzattoNo ratings yet
- 3-Ascending Tracts of Spinal CordDocument28 pages3-Ascending Tracts of Spinal Cordmuhammad altaf100% (2)
- How To Read A Head CT ScanDocument60 pagesHow To Read A Head CT ScanPompomNo ratings yet
- Types of Cerebral Herniation and ImagingDocument13 pagesTypes of Cerebral Herniation and ImagingPaulo LuizNo ratings yet
- Chapter One A CellDocument46 pagesChapter One A CellLeon MarkoNo ratings yet
- Glial CaellsDocument4 pagesGlial CaellsEmerson BartleyNo ratings yet
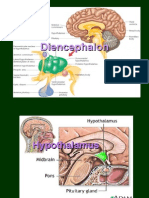
- HypothalamusDocument39 pagesHypothalamusAfsaneh Javanmard100% (1)
- Psycholinguistics: Language Disorder: Broca's AphasiaDocument18 pagesPsycholinguistics: Language Disorder: Broca's AphasiaSuciati AnandesNo ratings yet
- Psych Ass ReviewerDocument3 pagesPsych Ass ReviewerPINEDA, Maria Fatima D.No ratings yet
- Basic Neuroanatomy and Stroke Syndromes PDFDocument15 pagesBasic Neuroanatomy and Stroke Syndromes PDFFrancisco A. Villegas-López100% (2)
- Chapter 13: Spinal CordDocument15 pagesChapter 13: Spinal CordDao TranNo ratings yet
- The Nervous SystemDocument25 pagesThe Nervous SystemRoan Angela100% (4)
- ScienceSLM G10 Q3 M3 the-Nervous-System - Docx-1Document31 pagesScienceSLM G10 Q3 M3 the-Nervous-System - Docx-1John100% (1)
- Gerinlect2401 - 002lab104f16Document13 pagesGerinlect2401 - 002lab104f16api-282601291No ratings yet
- PiXL Knowledge Test ANSWERS - AQA B1 CORE Science - Legacy (2016 and 2017)Document12 pagesPiXL Knowledge Test ANSWERS - AQA B1 CORE Science - Legacy (2016 and 2017)Mrs S BakerNo ratings yet
- Quiz #1 Q3 Science 10Document2 pagesQuiz #1 Q3 Science 10Jr Capanang0% (1)
- Structures of The Central and Peripheral Nervous Systems WorksheetDocument3 pagesStructures of The Central and Peripheral Nervous Systems Worksheetapi-508202933No ratings yet
- Neuroscience NotesDocument4 pagesNeuroscience NotesAijia HeNo ratings yet
- Invertebrate Nervous Systems: Thomas MathesonDocument6 pagesInvertebrate Nervous Systems: Thomas MathesonMax MoralesNo ratings yet
- Anatomy and Physiology I (BIO201) Course MaterialsDocument18 pagesAnatomy and Physiology I (BIO201) Course MaterialsAnnie Minnie0% (1)
- Language and The BrainDocument8 pagesLanguage and The BrainAntonio GoqueNo ratings yet
- The Nervous System by MR RosenDocument10 pagesThe Nervous System by MR Rosenapi-202513022No ratings yet
- Frog Nervous SystemDocument24 pagesFrog Nervous SystemBeaNo ratings yet
- The CerebrumDocument27 pagesThe Cerebrumapi-19916399No ratings yet
- CLASS 10 Science Worksheet (2023-24)Document4 pagesCLASS 10 Science Worksheet (2023-24)SatyamNo ratings yet
- Lennart Heimer M.D. (Auth.) - The Human Brain and Spinal Cord - Functional Neuroanatomy and Dissection Guide-Springer-Verlag New York (1983)Document395 pagesLennart Heimer M.D. (Auth.) - The Human Brain and Spinal Cord - Functional Neuroanatomy and Dissection Guide-Springer-Verlag New York (1983)αργυ αργυ100% (1)
- Chapter - 005 Interdependent Body SystemsDocument46 pagesChapter - 005 Interdependent Body SystemsTina VargheseNo ratings yet
- Science: Quarter 2 - Module 2: The Human Body SystemsDocument32 pagesScience: Quarter 2 - Module 2: The Human Body SystemsJonel Buergo100% (9)
- VOL 3 - Layout 1Document596 pagesVOL 3 - Layout 1Raluca Crina100% (1)