Professional Documents
Culture Documents
Curs de Neuropsihologie
Uploaded by
papusica_mica0 ratings0% found this document useful (0 votes)
33 views63 pagesNeuropsihologie
Copyright
© Attribution Non-Commercial (BY-NC)
Available Formats
PDF, TXT or read online from Scribd
Share this document
Did you find this document useful?
Is this content inappropriate?
Report this DocumentNeuropsihologie
Copyright:
Attribution Non-Commercial (BY-NC)
Available Formats
Download as PDF, TXT or read online from Scribd
0 ratings0% found this document useful (0 votes)
33 views63 pagesCurs de Neuropsihologie
Uploaded by
papusica_micaNeuropsihologie
Copyright:
Attribution Non-Commercial (BY-NC)
Available Formats
Download as PDF, TXT or read online from Scribd
You are on page 1of 63
1 Curs de neuropsihologie
Universitatea din Bucureti
Facultatea de Psihologie i tiinele Educaiei
Andrei Chiri
2008
CURS DE NEUROPSIHOLOGIE
Cuprins
Cuvnt nainte ....................................................... 3
1. Relaia psihic-creier ........................................ 5
2. Structura microscopic i biochimia
sistemului nervos ................................................. 8
2.1. Neuronul ............................................................ 8
2.2. Celulele gliale .................................................. 10
2.3. Potenialul de membran celular ........... 10
2.4. Neurotransmitorii ..................................... 11
3. Anatomia funcional a sistemului
nervos central uman ........................................ 14
3.1. Mduva spinrii ............................................. 16
3.2. Trunchiul cerebral ........................................ 18
3.2.1. Bulbul rahidian (medulla oblongata) ...... 20
3.2.2. Pons (puntea lui Varolio) .............................. 22
3.2.3. Mezencefalul (creierul mijlociu)................ 22
3.2.4. Formaiunea reticulat (FR) ........................ 24
3.2.5. Patologia trunchiului cerebral .................... 25
3.2.6. O divizare alternativ i histochimia
trunchiului cerebral....................................................... 26
3.2.7. Nervii cranieni ..................................................... 27
3.3. Cerebelul .......................................................... 29
3.4. Diencefalul ....................................................... 32
3.4.1. Talamusul............................................................... 33
3.4.2. Metatalamusul ..................................................... 35
3.4.3. Epitalamusul ......................................................... 35
3.4.4. Subtalamusul ........................................................ 36
3.4.5. Hipotalamusul ..................................................... 36
3.5.Telencefalul ..................................................... 44
3.5.1. Sistemul (lobul) limbic .................................... 48
3.5.2. Nucleii bazali ........................................................ 52
3.5.3. Scoara cerebral ............................................... 55
4. Metode i tehnici de investigare n
neuropsihologie .................................................57
Bibliografie ..........................................................63
3 Curs de neuropsihologie
Cuvnt nainte
Nu exist fum fr foc si nici activitate uman
Ir motivatie. Asa c, nainte de a ncepe studiul
cursului de fat, am putea (si chiar ar trebui) s ne
ntrebm care este motivul care st la baza acestei
activitti. La prima vedere, rspunsul nu este greu de
gsit, acesta aflndu-se n examenul pe care va trebui
s-l absolvim. Apoi, dintr-un punct de vedere
anticipativ, absolvirea examenului constituie, la rndul
su, un motiv pentru un alt scop, respectiv obtinerea
licentei, care va conduce la obtinerea unui post, care
reprezint o surs de venit, asigurndu-ne astfel un trai
s.a.m.d. Desigur, acestea sunt niste motive generale, dar
fiecare dintre noi are, pe lng ele, si altele, particulare,
subiective.
Din pcate, foarte putini sunt cei ce se gndesc
dac poate exista un motiv intrinsec pentru a nvta
ceva, mai ales dac e vorba de un curs de
neuropsihologie. Un curs cum ar fi, de pild, cel de
psihologie social, cel de psihopatologie sau cel de
psihanaliz, este ntr-adevr atractiv pentru un student
la psihologie, deoarece expune niste cercetri cu o mare
aplicabilitate n viata de zi cu zi, ofer rspunsuri unor
ntrebri pe care si le-a pus studentul n legtur cu
anumite observatii empirice, sap adnc psihicul
uman, scotnd la iveal niste lucruri uneori aproape
mistice (de multe ori am auzit c inconstientul este un
trm, dar nu de la studenti).
ns, dup cum observ si Kolb si Whishaw
(2003), pentru studentul nceptor, nomenclatura
[totalitatea termenilor specifici] pentru nucleii si
tracturile sistemului nervos poate prea haotic. Asa
este. Multe structuri au mai multe nume, fiind folosite
de multe ori alternativ (p. 50). Spre exemplu, dac nu
era de ajuns c bulbul rahidian este un termen nu
tocmai comun, se mai face referire la acesta si cu
termenul de medulla oblongata ! Sau, nu era de ajuns
denumirea de vestibulocerebel, mai trebuia s se
numeasc si lob flocculonodular sau arhicerebel !
Aceast diversitate de termeni se datoreaz istoriei
lungi si complexe a neurostiintelor (ibidem). Termenii
folositi de anatomisti sunt mprumutati din greac,
latin, francez s.a. Totusi, majoritatea acestor termeni
sunt tradusi n romn, iar studentul trebuie s retin
doar una dintre denumiri, la alegere. Personal, mi se
pare c sunt mai usor de retinut termenii carenu sunt n
limba romn.
Pe lng acestea, imaginatia neuroana-
tomistilor compar structurile cerebrale cu anatomia
corpului (corpi mamilari), cu flora (amigdala sau
almond), cu fauna (hipocamp, sau clut de mare) si
cu personajele mitologice (cornul lui Ammon, care este
o parte a formatiunii hipocampice). O parte din
terminologie este un tribut adus anatomistilor pionieri :
fasciculul Vicq dAzyr, aria Broca, Wernicke, scizura
lui Rolando etc. Alti termeni se bazeaz pe culori :
substanta neagr (substantia nigra), aria albastr (locul
coeruleus) si nucleul rosu. Cel mai lung nume pentru o
structur cerebral este nucleus reticularis tegmenti
pontis Bechterewi, prescurtat NRPT, deoarece, dup
cum veti putea observa, oamenii de stiint au o afinitate
pentru abrevieri. Unele denumiri descriu consistenta :
substanta gelatinoas ; altele lipsa de cunostinte : zona
incert, nucleul ambiguu. Unele sunt bazate integral pe
o form ct mai scurt : grupurile de celule de la A-1 la
A-9 sau de la B1 la B9 (ibidem).
Toate acestea fac dificil, uneori imposibil,
plcerea de a nvta neuropsihologie. Dac, ns, mai
exist o sperant, atunci aceasta rezid n talentul
profesorului si al autorilor de manuale (tratate) de a
explica pe ntelesul tuturor si de a elimina multe dintre
elementele nesemnificative. Tratatul din care am citat
mai sus (Kolb, B, Whishaw, IQ, 2003, Fundamentals of
Human Neuropsychology, 5
th
edition) mi se pare a fi
foarte potrivit pentru aceast disciplin, deoarece
ndeplineste cele dou conditii de care vorbeam (si nu
numai). Un altul, la fel de bun, pare a fi : Carlson, N
(2005), Foundations of Physiological Psychology, 6th
ed., dar si Brain Facts A Primer on the Brain and
Nervous System, 4
th
ed., editat de Societatea Pentru
Neurostiinte. Din pcate, aceste crti nu pot fi
comandate dect pe internet, la niste preturi destul de
inaccesibile. Singurul tratat romnesc de neuropsiho-
logie de pe piat (din cte stiu eu) este cel de la Editura
Medical (Dnil, Golu, 2006), un tratat aproape
enciclopedic, dar care, din pcate, se adreseaz mai
degrab specialistilor dect studentilor si este plin de
greseli de redactare. De recomandat pare a fi si
compendiul de anatomia si fiziologia omului (Niculescu
et al., 2007, Editura Corint), dar este mai greu de gsit
(probabil doar prin anticariate).
Cursul de fat a fost realizat minutios, fiind
studiate mai multe surse pentru fiecare amnunt.
Deoarece nu este o lucrare proprie, ci mai degrab o
transcriere revizuit si adugit a cursului predat la
facultate, nu am putut s urmez un alt plan, care poate
ar fi atras mai mult cititorul. Din pcate, s-a insistat
prea mult pe anatomia functional a fiecrei structuri
cerebrale si, de aceea, nu au mai fost prezentate
principalele sisteme functionale (senzorial, motor,
emotional, intelectuale etc.). Astfel, cursul a devenit,
mai degrab de anatomie si, partial, fiziologie,
ignorndu-se destul de mult faptul c totusi e vorba de
un curs de psihologie.
Imaginile pe care le-am introdus n acest curs
sunt de maxim important, fiind, de altfel, motivul
pentru care am decis s transcriu cursul. De aceea, nu
pot oferi un sfat mai bun cititorului dect acesta : nu
trece mai departe pn nu ai nteles o imagine ! Dup
citirea textului (sau n paralel), ntelegerea imaginilor
nu este dificil si probabil nu va dura mai mult de 5-10
minute. Si nu exist metod mai bun de a nvta
anatomie dect prin imagini. Cine doreste si are timp
poate s le deseneze cu mna sa, lucru de asemenea
foarte util. Din motive de spatiu (megabiti), imaginile
au fost comprimate si, din pcate, nu au o calitate foarte
bun. Am ncrederea c acest document sau imaginile
din el nu vor aprea prin alte locuri de pe internet !
Pe lng imagini, am introdus si cteva tabele
(mai putine dect as fi dorit), utile pentru referinte
rapide. n redactare am ales modelul mprtirii paginii
n dou coloane, format din ce n ce mai folosit n
tratate si care face mai comod lectura. Unele din
unittile pe care le-am introdus n plus fat de ce a fost
predat la curs faciliteaz ntelegerea (cum ar fi cap. 2.3.
Potentialul de membran celular), iar altele sunt pur si
simplu optionale, pentru cine vrea s stie mai mult (cum
ar fi alte functii ale hipotalamusului). Aceasta nu
nseamn, ns, c cele optionale nu faciliteaz, ntr-o
msur mai mic, ntelegerea. Tot ce este n plus a fost
notat. S-au strecurat, cu sigurant, greseli, dar sper ca
ele s nu fie de continut, ci doar de redactare. n orice
caz, documentul va fi revzut.
Un alt sfat util ar fi acela de a nu se ncerca
nvtarea dintr-o dat. Pentru integrarea informatiilor
este absolut necesar trecerea a cel putin de dou ori
peste fiecare unitate, eventual dup anumite perioade de
timp. Cantitatea de informatii este considerabil, iar
ncercarea de a memora pe de rost este futil. Cele
patru situri de la sfrsitul bibliografiei se pot dovedi
foarte utile pentru cei care nu au un atlas. La adresa
http://www.neuroanatomy.wisc.edu/, sectiunea
Neuroscience video demonstrations se pot gsi cteva
videoclipuri care prezint disectia trunchiului cerebral.
Cu speranta c acest curs se va dovedi folositor,
v urez rbdare n parcurgerea lui si succes la examenul
care va urma.
Colegul vostru,
Andrei Chirit.
5 Curs de neuropsihologie
1. RELAIA PSIHIC-CREIER
Relatia dintre psihic i creier reprezint
obiectul de studiu al neuropsihologiei. Neuropsihologia
studiaz cum modificrile de la nivelul creierului afec-
teaz comportamentul : cum, de exemplu, ablatia (extir-
parea) regiunilor prefrontale ale creierului, la om, i va
modifica acestuia capacittile intelectuale sau
caracterul, cum ablatia regiunilor infero-temporale ale
creierului maimutei va influenta perceptia si memoria
vizual, cum ablatia unor portiuni din cortexul cerebral
la sobolani va modifica nvtarea unui labirinit cu mai
multe locuri nfundate.
Desigur, aceste exemple se refer la expe-
rimente de laborator, dar neuropsihologia poate studia
si alte fenomene, produse natural. De exemplu, la
nceputul capitolului Bazele biologice ale psihologiei
dinIntroducerea n psihologie Atkinson & Hilgard, ed.
a XIV-a, este prezentat cazul unui angajat al cilor
ferate, Phineas Gage, care, n 1848, a suferit un
accident oribil si a rmas numai o umbr a celui care
fusese pn atunci . Un ax de fier cu lungimea de peste
un metru a ptruns, n urma unei explozii, prin obrazul
stng, apoi prin spatele ochiului stng si a iesit din
craniu prin crestetul capului lui Gage, fiind gsit la
ctiva metri de angajat, acoperit de snge si resturi de
creier. Gage a supravietuit, n chip miraculos, iar dup
mai bine de o lun de trud a reusit s se ntoarc acas.
ns, dintr-un brbat blnd si rezervat, Gage ajunsese
acum zgomotos si impulsiv, cu preferinte pentru lucruri
vulgare. A ajuns, pn la urm, un vagabond si a murit
n 1860 de epilepsie.
n general, n neuropsihologie se diferentiaz :
neuropsihologia clinic, neuropsihologia experimental,
neurologia comportamentului si neuropsihologia cogni-
tiv.
Evidentierea si afirmarea legturii dintre cele
dou entitti (psihic si creier) se realizeaz abia n anti-
chitatea trzie, doar cu cteva secole nainte erei noastre
si mult mai trziu dect aparitia constientizrii si anali-
zrii psihicului. n sec. V .e.n., Hippocrate(cca. 460
cca. 370 .e.n.) si Kroton considerau c creierul este
sediul gndirii, ratiunii, n general, a proceselor
cognitive intelectuale, iar procesele afective erau
localizate n inim. Putem observa c aceast conceptie
este cea mai popular, specific, de regul, simtului
comun (n-ai inim!, ce-mi trece prin cap etc.), dar
asta nu nseamn c erau departe de adevr, n special
n ce priveste functiile intelectuale.
Mai trziu, Galen (129 200/216) formuleaz
prima ipotez despre o localizare direct a functiilor si
proceselor psihice n structurile cerebrale. El considera
c impresiile din lumea extern ptrund, n forma
fluidelor, prin ochi, n ventriculii cerebrali, unde se
grupeaz cu lichidele vitale sosite din ficat, transfor-
mndu-se n fluide psihice (pneuma psihikon sau
pneuma loghistikon). Ideea a fost revolutionar pentru
vremea respectiv, dar, desigur, astzi este naiv. Cre-
dinta c lichidul care irig ventriculii cerebrali consti-
tuie substratul material nemijlocit al psihicului a domi-
nat peste un mileniu si jumtate !
n sec. XVII, Ren Descartes (1596-1650)
avanseaz ipoteza c ntregul nostru psihic este situat n
glanda epifiz, situat central la baza emisferelor
cerebrale, pozitie care-i confer, n opinia lui, rolul de
dispecer al spiritelor animale, purttoarele psihicului.
ncepnd cu anatomistul german Meyer (1779),
apare bine postulat ideea localizrii distincte a proce-
selor psihice, adic fiecare functie psihic putea fi gsi-
t ntr-o anumit parte a creierului. Ideea a fost contura-
t mai bine de Franz J osef Gall (1758-1828), care
consider c n spirit exist facultti separate (inteli-
genta, memoria, perceptia etc.), iar fiecare din aceste
facultti ar avea localizri precise n anumite regiuni ale
creierului. Gall considera c oasele cutiei craniene se
vor dezvolta n raport cu nivelul de dezvoltare al
cortexului si este suficient palparea craniului pentru a
examina cortexul. Metoda lui s-a numit initial
cranioscopie, apoi frenologie. Ideea lui Gall atrage
atentia, pentru prima dat, asupra caracterului
diferentiat al scoartei cerebrale care nu mai este un
simplu conglomerat de neuroni, iar ideea sa a
directionat mai bine de 100 de ani de cercetri
localizaioniste asupra creierului.
Cercetrile clinice ale lui Paul Broca (1824
1880) si ale lui Carl Wernicke(18481904) au condus
la primele descoperiri concrete asupra legturii dintre
anumite arii (ce le poart numele) si functia psihic a
limbajului.
Broca descoper c, la pacientii si cu
deficiente grave de vorbire, era lezat portiunea
posterioar (piciorul) a circumvolutiunii frontale infe-
rioare din emisfera stng. A numit aceast tulburare
afazie motorie.
Wernicke, n 1871, descoper c pacientii si
aveau lezat circumvolutiunea temporal superioar tot
6 Curs de neuropsihologie
din emisfera stng. Respectivii pacienti nu putea nte-
lege limbajul oral (afazie senzorial).
De acum ncolo, avem de-a face cu o adev-
rat explozie de studii localizationiste. Cercetrile lui
Betz (1874) asupra neuronilor piramidali giganti (care i
poart numele) asociati cu miscarea, experimentele lui
Munk (1881) asupra recunoasterii vizuale, prin ex-
tirparea unor portiuni din lobii occipitali, precum si alte
multe cercetri, l-au condus pe Vogt (1951) la realiza-
rea unui model topic al organizrii functionale a creie-
rului. n timpul celui de-al doilea rzboi mondial, avnd
acces la examinarea rnitilor, Luria (1947) ntreste
ideea c orice functie psihic se leag de anumite struc-
turi sau formatiuni cerebrale, dar nu accept ideea unei
localizri precise, punctiforme, drept care formuleaz
ipoteza localizrii dinamice. Luria nu respinge ideea c
unele functii psihice, cum ar fi cele senzoriale, au o
localizare precis, dar postuleaz c cele superioare nu
au o localizare precis, ci depind de mai multe forma-
tiuni cerebrale.
Trebuie s retinem c modelul neuroatanomic
vizeaz att structurile si formatiunile anatomice n
sine, ct si legturile dintre acestea, care sunt la fel de
importante, lezarea lor ducnd la efecte psihocomporta-
mentale specifice.
Gazzaniga si Sperry (1967, 1970) au demon-
strat, pentru prima dat, specializarea functional a
celor dou emisfere cerebrale. Modelul creat de cei doi
poart numele desplit-brain (creier divizat).
n concluzie, orientarea neuroanatomic locali-
zationist sustine urmtoarele :
Fiecare functie psihic are o reprezentare cerebral
separat ;
Centrele corticale se leag ntre ele prin fascicule
de substant alb (nervi) ;
Efectele neuropsihopatologice variaz n functie de
lezarea centrilor, a substantei albe sau a ambelor.
Modelul localizationist nu a putut dobndi o
recunoastere unanim. Dac datele si argumentele sale
referitoare la localizarea functiilor psihice simple
(senzoriale si motorii) erau suficient de convingtoare,
cele referitoare la localizarea functiilor psihice supe-
rioare (gndire, memorie, imaginatie, motivatie, voint)
strneau nedumerire si ndoial.
Astfel, ncepe disputa dintre Pierre Flourens
(17941867) si Gall. Am vzut modelul localizationist
al lui Gall. Flourens observ, prin extirparea unor
portiuni cerebrale la porumbel, c, dup trecerea unui
anumit interval de timp, tulburrile aprute initial se
diminueaz sau chiar dispar. Aceasta se explic prin
faptul c o alt portiune cerebral poate prelua functiile
celei extirpate (compensare). Fenomenul compensrii
se desfsura la fel, indiferent de portiunea extirpat.
Modelul lui Flourens se numesteechipotenialism. Spre
deosebire de localizationismul lui Gall, care era pur
speculativ, echipotentialismul lui Flourens se bazeaz
pe date empirice, experimentale. Flourens face totusi
greseala de a extrapola datele obtinute la porumbel si
asupra omului, nelund n considerare primitivitatea
creierului de porumbel n comparatie cu cel uman.
Prin urmare, echipotentialismul afirm c
scoarta cerebral functioneaz ca un tot amorf,
nediferentiat, n ciuda complexittii ei, iar lezarea
diferitelor portiuni poate provoca tulburri, att n
sfera senzorial, ct i n cea intelectual. Desigur, ca
si n cazul localizationismului, echipotentialismul a dat
nastere la numeroase cercetri n urmtorii zeci de ani.
n sprijinul echipotentialismului aduce dovezi
Goltz (ntre 1876 si 1881), iar mai trziu, Kark Lashley
(1929). Acesta efectueaz experimente pe cobai, crora
le extirpa portiuni de diverse mrimi din creier si le
urmrea evolutia tabloului comportamental n sarcina
labirintului. n primele zile dup operatie, animalele
erau dezorientate si nu se puteae orienta n spatiu, pre-
zentnd si tulburri de coordonare motorie. Treptat,
aceste tulburri se diminuau, n final, comportamentul
ajungnd la un nivel de eficient bun. Lashley a consta-
tat c gradul si durata de realizare a compensrii depin-
deau de mrimea suprafetei extirpate. Pe baza acestor
date, Lashley realizeaz forma complet a modelului
echipotentialist clasic, postulnd urmtoarele :
Nu exist o legtur direct si stabil (predetermi-
nat) ntre natura tulburrii si locul leziunii
cerebrale ;
n procedura tulburrilor functionale, esential este
ntinderea zonei lezate si nu localizarea ei ;
Tulburrile functionale provocate de leziuni sau
focare limitate ale creierului au caracter tranzitoriu,
ele fiind compensate prin prelucrarea functiei de
ctre alte zone, rmase integre.
n plan psihologic, modelul ngust localizatio-
nist ia ca suport teoria asociationist, iar cel echipo-
tentialist teoria gestaltist.
Evolutia ulterioar a neuropsihologiei a
demonstrat c, att localizationismul ngust, ct si
echipotentialismul nu ofer rspunsuri corecte si
exhaustive la toate ntrebrile pe care le ridic, n mod
concret, raportul psihic-creier. J ohn Hughlings J ackson
(18351911) si Ivan Petrovici Pavlov (1849-1936) au
impus perspectiva genetic-evoluionist n ntelegerea
relatiei dintre psihic si creier. Aceast perspectiv arat
c mecanismul neuronal al unei functii psihice nu este
7 Curs de neuropsihologie
nnscut, predeterminat, ci se constituie n cursul
evolutiei filo- si ontogenetice, o dat cu si pe msura
aparitiei si dezvoltrii functiei nsesi. Din acest punct de
vedere, nici o functie psihic nu trebuie privit ca un dat
si nici raportat la o structur anume, care, n sine, este
inert. Perspectiva genetic-evolutionist sustine c
functia se integreaz n structur, ambele formnd o
unitate dinamic evolutiv.
Fundamentul acestei perspective se bazeaz pe
constatarea clinic a lui Jackson c o functie psiho-
logic nu se pierde niciodat complet n urma unei
leziuni cerebrale. Constituirea functiilor psihice, arat
J ackson, se va efectua dup schema tripl care urmeaz:
1) evolutia este trecerea de la mai putin organizat la
organizat ; 2) evolutia este trecerea de la simplu la
complex ; 3) evolutia este trecerea de la automat la
voluntar.
Mai mult, functiile psihice nu pot fi suprapuse
n mod nemijlocit peste structura anatomic a
creierului, adic, de exemplu, un creier nu gndeste prin
simplul fapt c exist. Rolul principal al functionrii
psihice revine dinamicii corticale, proceselor
fundamentale ale acesteia (excitatia si inhibitia). Deci,
functiile psihice apar si se manifest ca rezultat al
interactiunii dintre excitatie si inhibitie, interactiune
care se desfsoar pe un spatiu ntins, cuprinznd un
numr mare de puncte si zone neuronale. Pavlov a
introdus termenul de mozaic, care reprezint, ntr-o
manier plastic, dinamica cortical. Mozaicul
reprezint desfsurarea unui anumit proces psihic prin
modificarea permanent a tabloului electric (tabloul
activismului bioelectric). Prin urmare, dac am putea
vizualiza scoarta cerebral, aceasta ar arta ca un
mozaic, plin de puncte luminoase care se sting si se
aprind extrem de rapid, lucru care, de altfel, a fost
demonstrat cu tehnicile de imagistic cortical.
Anohin (1935, 1940), apreciind ideile lui
J ackson, elaboreaz conceptul desistem functional, care
permite o mai bun ntelegerea a integrrii
neuropsihologice. Functia psihic devine, la Anohin, un
instrument de adaptare a individului la mediu. Din acest
punct de vedere, functia psihic trebuie considerat ca
un lant de operatii si verigi legate ntre ele pe baza
efectului adaptativ. Mai trziu, Bernstein (1947) a
demonstrat pe larg caracterul sistemic-ierarhizat al
actelor motorii. Fr a intra n detalii, precizm c orice
act motor se constituie n raport cu anumite sarcini si
solicitri.
Moruzzi si Magoun (1949) au artat, pe baza
cercetrilor asupra formatiunii reticulate, c o functie
psihic include si mecanisme nespecifice, subcorticale
(cum este, desigur, chiar formatiunea reticulat, vezi
cap. 3.2.4.). Astzi, neuropsihologia concepe realizarea
oricrei functii psihice ca interactiune ntre structurile
neuronale specifice, corticale si cele nespecificice,
subcorticale.
Aleksandr Romanovich Luria (1902-1977), n
lucrrile sale de dup anul 1960, a dezvoltat si
argumentat schema logic i operaional a
localizrilor dinamice. El a artat c, n problema
raportului psihic-creier, trebuie s distingem un aspect
fundamental si unul secundar.
Aspectul fundamental se refer la faptul c nici
un proces psihic nu poate apare n afara creierului sau
n afara functionrii lui sub influenta unor informatii
exterioare. Prin urmare, organizarea psihic, n
ansamblu, trebuie interpretat ca expresie si rezultat al
activittii reflexe a creierului ca sistem, aceasta
presupunnd admiterea legturilor si interactiunilor, att
pe vertical, ct si pe orizontal.
Aspectul secundar ridic probleme de gsire a
mecanismului prin care se realizeaz un proces psihic si
de determinare a nivelului anatomic la care se
integreaz o functie psihic. Pentru acestea, trebuie s
tinem cont de anumite aspecte, cum ar fi : succesiunea
formrii functiilor psihice si a structurilor neuronale n
filo- si ontogenez, gradul de complexitate al functiilor
psihice, plasticitatea functional a structurilor cerebrale
si gradele de libertate combinatoric proprii neuronilor
ce alctuiesc diferite structuri si zone ale creierului.
Astfel, ajungem s delimitm n interiorul sistemului
nervos central (SNC) dou mari tipuri de structuri :
specializate, nchise (cum ar fi aria Broca, Wernicke,
ariile vizuale etc.) si nespecializate, deschise (structuri
alocate limbajului, gndirii, imaginatiei, motivatiei,
emotiei etc.). De aici, se pot mprti si functiile psihice
n : functii cu localizare precis si invariant (functiile
senzoriale si motorii) si functii cu localizare relativ,
dobndit (restul).
Cercetrile ntreprinse dup anii 60 (Eccles,
Borgen, Pribram, Botez) au demonstrat caracterul
modular al organizrii structural-functionale a scoartei
cerebrale. Anatomic, modulul este un subansamblu de
aproximativ 10.000 de neuroni dispusi n coloane, n
interiorul crora se stabilesc complexe raporturi func-
tionale ntreinput-urile excitatorii si cele inhibitorii. In-
tegrarea functiilor psihice se face secvential, de la ariile
primare la cele secundare, iar de la acestea, la cele
tertiare si quternare (supraordonate).
8 Curs de neuropsihologie
2. STRUCTURA MICROSCOPIC I BIOCHIMIA
SISTEMULUI NERVOS
Structura general a SN cuprinde trei compo-
nente principale : 1) un compartiment senzorial (recep-
torii senzoriali) ; 2) un compartiment motor (efectorii) ;
3) un compartiment asociativ-integrativ (ariile de aso-
ciatie, corticale si subcorticale).
Semnalele senzoriale venite pe calea organelor
de receptie (ochi, urechi, limb etc.) sunt transmise
ctre zona de asociatie, unde pot crea o reactie imediat
sau pot fi memorate pentru un timp de ordinul minute-
lor, sptmnilor, anilor, putnd apoi ajuta la conditio-
narea reactiilor organismului ntr-un moment viitor.
99% din informatiile senzoriale cu care este bombar-
dat creierul sunt eliminate ca fiind nesemnificative sau
neimportante.
SNC uman a mostenit capacitti functionale
speciale din fiecare etap evolutiv a dezvoltrii. Exist
trei niveluri principale ale SNC, fiecare cu caracteristici
functionale specifice : 1) nivelul mduvei spinrii ; 2)
nivelul subcortical (cerebral inferior) ; 3) nivelul corti-
cal (cerebral superior). Cele dou proprietti fundamen-
tale ale organismelor vii, excitabilitatea (capacitatea de
a rspunde la un stimul, prin manisfestri caracteristice)
si conductibilitatea (capacitatea de a transporta
semnale) ating cea mai mare dezvoltare la SNC uman.
Unitatea de baz a SN este neuronul, o celul
specializat care transmite impulsuri neuronale (mesaje)
altor neuroni, glande sau muschi. Neuronii detin secre-
tul functionrii creierului si sunt responsabili pentru
existenta constiintei. Neuronii rspund la stimuli, trans-
port semnale i proceseaz informatie. Aceste trei sar-
cini fac posibil existenta proceselor psihice. n SN se
gsesc neuroni dispusi n serii longitudinale sau n
legturi de tip serial. Aranjamentul serial formeaz
dou tipuri de circuite : reflex (transport impulsuri care
conduc la un rspuns involuntar la un stimul ; de exem-
plu, reflexul rotulian) si de releu (circuitul somatic
aferent general, care transport impulsuri de la o parte
la alta a SN, cum este, de exemplu, transmiterea infor-
matiei senzoriale la creier si formarea senzatiei).
Circuitele reflexe se pot suprapune cu prti ale circuite-
lor somatice aferente (de releu). Semnalele ce sosesc
ctre SNC sunt numiteaferente, iar cele care pleac de
la SNC ctre muschi si organele efectoare se numesc
eferente.
2.1. NEURONUL
Neuronii sunt celule care actioneaz ca unitti de
transmitere a informatiei n interiorul SN si, desi au
multe caracteristici n comun cu alte celule din corp,
prezint si caracteristici speciale care le permit s nde-
plineasc sarcina de transmitere a informatiei. Dup
functia lor general, neuronii se mpart n neuroni
senzitivi (transmit impulsurile nervoase de la receptori
la SNC), neuroni motori (transmit semnalele eferente
de la SNC la muschi si glande) si neuroni de asociatie
sau intercalari (primesc semnalele de la neuronii
senzitivi si le transmit altor neuroni). Desi variaz
considerabil ca form si mrime, ei au caracteristici
comune. Un neuron tipic este format din corp celular
(soma) si procesele protoplasmatice (dendrite si un
axon) (fig. 2.1).
Corpul celular este centrul metabolic al neuro-
nului si contine nucleul si citoplasma. n SNC, un grup
format din mai multe corpuri celulare se numeste
nucleu, iar n SNP ganglion.
Nucleul este cel mai important element al neu-
ronului, dar, dup completa dezvoltare a SN, acesta, din
pcate, nu sufer mitoze nu se poate reproduce.
Nucleul contine nucleoplasma, un pigment numit
cromatin, un nucleol proeminent si, doar la femeie, un
satelit nucleolar.
9 Curs de neuropsihologie
Citoplasma contine organitele celulare obisnui-
te : mitocondriile, aparatul Golgi, lizozomii, reticulul
endoplasmatic si corpusculi Nissl.
Dendritele sunt scurte si transport impulsul
nervos ctre corpul celular.
Axonii nu contin corpusculi Nissl, variaz n
lungime de la micrometri la metri si transport impul-
suri nervoase de la soma ctre periferie. Axonii sunt
singurele componente ale neuronilor care se pot
regenera, dar aceastanumai n SNP, n SNC nici mcar
acestia neavnd posibilitatea de regenerare. n interiorul
unui axon sunt prezente dou tipuri de fluxuri de lichide
(axoplasmice) : fluxul axoplasmic anterograd (de la
soma la periferie) si fluxul axoplasmic retrograd (de la
periferie ctre soma). Transportul axoplasmic antero-
grad este vital pentru cresterea axonului n cursul dez-
voltrii, pentru mentinerea structurii axonale si pentru
sinteza si eliberarea neurotransmittorilor. Transportul
axoplasmic retrograd are important clinic, fiind calea
pe care intr toxinele si virusurile n SNC. La captul
unui axon se gsesc niste terminatii, numite si butoni
terminali, prin intermediul crora se realizeaz sinapsa
cu urmtorul neuron, cu un muschi sau cu o gland.
Unnerv este un fascicul format din axoni lungi
care apartin mai multor sute de neuroni. Un singur nerv
poate contine att axoni ai neuronilor senzitivi, ct si
neuroni ai neuronilor motori.
Mielina este o substant gras, de culoare alb
(un fosfolipid) care acoper axonii unor anumiti neuroni
pe care i izoleaz electric, crescnd astfel viteza de
transmitere a impulsului nervos. Mielina este dispus
de-a lungul unui axon sub forma mai multor straturi
cu ct mai multe, cu att mai bine. Nu este un produs al
celulelor nervoase, ci al unor celule de sustinere
(oligodendrocite n SNC si Schwann n SNP ; vezi si
subtitlul urmtor).
n functie de procesele protoplasmatice,
neuronii pot fi : a) unipolari : contin doar axonul, fr
dendrite si se gsesc aproape numai n ganglionii
spinali si pe post de receptori n piele ; b) bipolari :
prezint un axon si o dendrit si sunt dispusi numai la
nivelul cilor auditive, vestibulare si vizuale ; c) multi-
polari : toate celelalte celule din SN sunt multipolare si
prezint de la una la 20 de dendrite (fiecare putnd avea
mai multe ramuri) si un axon.
Sinapsa este jonctiunea, legtura dintre un
buton terminal si un alt neuron (la nivelul corpului
celular, al dendritelor sau al altui axon), celul
muscular sau glandular (fig. 2.2). Un fapt important
este c ntre butonul terminal si urmtoarea celul
nervoas
1
exist un mic spatiu numit fant sinaptic.
Cnd un impuls neuronal coboar prin axon si ajunge la
butonii terminali, acesta declanseaz secretia unui
neurotransmitor, o substant chimic eliberat n
fanta sinaptic cu rolul de a stimula sau inhiba urmto-
rul neuron
2
. Sinapsele pot fi chimice (transferul se face
prin intermediul neurotransmittorilor) sauelectrice (cu
ajutorul unor canale apoase deschise care conduc
impulsul electric). Majoritatea sinapselor sunt chimice.
Procesele psihice sunt, practic, rezultatul sinapselor.
Polarizarea este principiul fiziologic al sinapsei
care face ca directia impulsului s fie ntotdeauna de la
axon ctre urmtorul neuron. Fiecare impuls poate fi :
blocat, transformat n impulsuri repetitive sau integrat
alturi de impulsurile provenite de la alti neuroni.
1
Se ntelege c aceleasi principii sunt valabile si pentru celelalte
dou tipuri de unitti anatomice cu care se poate face sinaps
(muschi, glande), pe care nu le vom mai aminti n continuare,
mentionnd doar sinapsele dintre neuroni.
2
Sinapsa dintre doi axoni (axoaxonal) nu are rolul de a conduce
impulsul nervos (stimula urmtorul neuron), ci modific volumul de
neurotransmittor eliberat de neuronul postsinaptic (n cadrul sinap-
sei axoaxonale).
Fig. 2.1. Schema neuronului. Neuron mielinizat din SNP
(deoarece are teaca de mielin Schwann, vezi cap. 2.2.)
Fig. 2.2. Schema sinapsei la nivelul corpului celular.
Curs de neuropsihologie 10
2.2. CELULELE GLIALE
O mare parte din celulele SN nu sunt neuroni, ci celule
care asigur protectia sau hrnirea (trofia) neuronilor.
Acestea sunt : a) celule osoase care formeaz cutia
cranian si coloana vertebral si b) celule de sustinere
(gliale). Celulele (neuro)glialenconjoar neuronii si se
afl n proportie de 10 pn la 50 de ori mare dect
neuronii din SNC al vertebratelor. La nivelul SNC,
deosebim trei categorii de celule gliale : ependimale,
microgliale si macrogliale.
Celulele ependimale tapeteaz cavittile pline
cu fluid (ventriculii cerebrali) si canalul central al m-
duvei spinrii.
Celulele microgliale sunt fagocite ce iau nastere
din macrofage si nglobeaz resturile rezultate din
lezarea, infectiile sau bolile SNC.
Macroglia este compus din patru tipuri de
celule : astrocite si oligodendrocite la nivelul SNC si
celule Schwann si capsulare la nivelul SNP.
Astrocitele sunt celule cu form de stea (astro-),
cele mai numeroase celule ale SNC. Astrocitele
formeaz un nvelis acoperitor numit membran
limitant extern saumembran glial. Procesele lor au
terminatii vasculare care nconjoar capilarele, formnd
bariera hemato-encefalic, ce permite trecerea selectiv
a substantelor din sngele circulant n SNC. Astrocitele
sunt cele mai afectate de iradierea SNC si sunt primele
care fac neoplasm (tumori) !
Oligodendrocitele sunt celule gliale a cror
functie principal este formarea si mentinerea mielinei
la nivelul SNC. Teaca de mielin se formeaz cu
ajutorul proceselor oligodendrocitelor, care se dispun n
jurul axonilor, formnd o spiral strns (fig. 2.2).
Oligodendrocitele pot mbrca, de asemenea, si corpii
neuronali dar, n acest caz, nu formeaz mielin. De
curnd, se presupune c oligodendrocitele au si un rol
de hrnire, ajutnd cresterea nervoas ce poate promova
cresterea axonilor afectati din SNC.
Celulele Schwann sunt corespondentele oligo-
dendrocitelor la nivelul SNP. Spre deosebire de
oligodendrocite, celulele Schwann nconjoar doar
partial axonul mielinizat. ntre fiecare celul Schwann,
mielina este ntrerupt, formnd ariile numite nodurile
lui Ranvier (fig. 2.1). Celulele Schwann au nc un rol
foarte important, si anume acela de regenerare a
axonilor distrusi (numai cei din SNP avnd aceast
capacitate si se presupune c nu se regenereaz si la
nivelul SNC tocmai datorit lipsei celulelor Schwann).
Celulele capsulare constituie elemente gliale ce
nconjoar corpii neuronilor la nivelul ganglionilor
senzoriali si autonomi.
2.3. POTENIALUL DE MEMBRAN CELULAR
(unitate nepredat la curs)
Dup cum se poate vedea n fig. 2.2, n cadrul unei
sinapse sunt implicate dou membrane : membrana
butonului terminal, care devinemembrana presinaptic
si membrana celulei nervoase, care devine membrana
postsinaptic. ntre cele dou suprafete (exterioar si
interioar) ale membranelor tuturor celulelor din
organism exist potentiale electrice. Neuronii sunt capa-
bili s genereze, la nivelul membranelor, propriile
impulsuri electrochimice care variaz rapid, acestea
fiind esentiale pentru transmiterea semnalelor de-a
lungul suprafetelor celulare. Pentru a ntelege mai bine,
putem compara neuronul cu o baterie. Aceast diferent
de potential este dat de distributia inegal a
concentratiei de ioni pe cele dou prti ale membranei,
fapt de mare important pentru functionarea celulelor
musculare si nervoase. Membrana plasmatic este un
perete viu, un nvelis care separ continutul celulei de
lichidul extern, intercelular ; mai exact, este o sit vie
care las s treac anumite substante, iar pe altele le
retine. Cile de acces de pe suprafata membranei se
numesc canale ionice si sunt niste molecule proteice
care formeaz pori. Fiecare canal ionic este selectiv,
permitnd unui singur tip de ion s treac prin el cnd
este deschis. Cei mai importanti ioni transportati de
aceste proteine sunt Na
+
, K
+
, Cl
-
, si Ca
2+
. Transportul
ionic celular nu ar avea nici o valoare dac fluxul n-ar fi
controlat si dac toate cele cteva mii de canale ionice
dintr-o membran ar fi deschise tot timpul.
Datorit distributiei inegale a diferitilor ioni n
mediul intra- si extracelular, se produce polarizarea
electric a membranei (un pol negativ n interior si unul
pozitiv n exterior). ntre cei doi poli apare o diferent
de potential, care poart denumirea de potenial de
repaus. La celulele nervoase ale vertebratelor, valoarea
11 Curs de neuropsihologie
medie a acestui potential este de -70 mV. n stare de
repaus, membrana celular nu este permeabil pentru
ionii de Na
+
si acestia se gsesc n concentratii mari n
afara neuronilor. n schimb, membrana este permeabil
fat de ionii de K
+
, care tind s se concentreze n inte-
riorul neuronului.
Mecanismul care asigur transportul activ al
ionilor de Na
+
n afar si al ionilor de K
+
din exterior n
interior n perioada de repaus poart numele depompa
Na
+
K
+
(pompa sodiu-potasiu). Pompa Na
+
K
+
este,
de fapt, o membran proteic si foloseste ca surs de e-
nergie acidul adenozintrifosfat (ATP), combustibilul
universal al tuturor celulelor (o molecul de ATP
permite expulzarea a 2-3 ioni de sodiu). ATP este
produs de ctremitocondrii (fig. 2.2).
O stimulare (n esent mecanic, chimic sau
electric) suficient de puternic determin cresterea
local a permeabilittii membranei, ceea ce are ca efect
un transfer energic de ioni dintr-o parte n alta. Ca
urmare a cresterii permeabilittii, diferenta de potential
dintre interiorul si exteriorul celulei se reduce, provo-
cnd o depolarizare electric. Ca urmare, neuronul se
descarc, dnd nastere unui potenial de aciune.
Potentialul de actiune este un impuls electrochimic,
nsusi impulsul nervos, care pleac din corpul celulei si
coboar pn la terminatiile axonului (butonii termi-
nali). Potentialul de actiune este crucial pentru propa-
garea la distant, fiecare neuron actionnd ca un nou
amplificator al acestui semnal, care si diminueaz in-
tensitatea datorit distantei. Depolarizarea se propag n
jos de-a lungul axonului, iar n urma trecerii potentialu-
lui de actiune, canalele de Na
+
se nchid n urma sa si
pompele ionice intr n actiune pentru a reduce mem-
brana celular la potentialul de repaus. Viteza potentia-
lului de actiune este de la 3 la 320 de km/h, n functie
de diametrul axonului axonii mai mari au, n general,
o propagare mai mare. Dup cum artam n cap. 2.1.,
viteza propagrii semnalului este facilitat de mielin.
Un singur neuron genereaz un potential de ac-
tiune cnd excitatia care ajunge la el prin multiple
sinapse depseste un anumit prag. Dac nu se atinge
acest prag minim, atunci neuronul nu creeaz un poten-
tial de actiune. Astfel, n urma informatiilor primite din
sinapse, neuronul emite sau nu un potential de actiune.
Aceast caracteristic a neuronului se numeste
principiul tot-sau-nimic. Din acest punct de vedere,
modul de functionare al SNC este asemntor cu codul
binar al calculatoarelor (0 si 1).
Dup cum artam n cap. 2.1., neuronii nu
realizeaz contact direct n cadrul sinapselor, semnalul
fiind nevoit s treac prin fanta sinaptic. Cnd un po-
tential de actiune coboar de-a lungul axonului si
ajunge la butonii terminali, acesta stimuleaz veziculele
sinaptice (fig. 2.2). Veziculele sinaptice sunt mici
structuri sferice care contin neurotransmittori, pe care
i elibereaz n fanta sinaptic la primirea impulsului
nervos. Neurotransmittorii se cupleaz cu receptorii,
niste proteine localizate n membrana postsinaptic.
Aceast cuplare declanseaz o schimbare direct n
permeabilitatea canalelor ionice n neuronul receptor.
Atunci cnd primesc un impuls, unii receptori
postsinaptici au rolul de a excita neuronul postinaptic,
iar altii de a-l inhiba.
Pentruexcitaie se produce : 1) deschiderea ca-
nalelor de sodiu pentru a permite ptrunderea unui nu-
mr mare de sarcini electrice pozitive n interiorul ce-
lulei postsinaptice, 2) scderea conducerii prin canalele
de clor sau potasiu, sau prin ambele si 3) diverse modi-
ficri ale metabolismului intern al neuronului postsinap-
tic, care conduc la stimularea activittii celulare sau, n
unele cazuri, la cresterea numrului de receptori mem-
branari excitatori sau scderea numrului de receptori
membranari inhibitori.
Pentruinhibiie se produce : 1) deschiderea ca-
nalelor ionice de clor din membrana postsinaptic ; 2)
cresterea conductantei ionilor de potasiu spre exteriorul
neuronului si 3) activarea unor receptori care inhib
functiile metabolice celulare, ceea ce conduce la
cresterea numrului de receptori inhibitori sau
reducerea numrului de receptori excitatori.
2.4. NEUROTRANSMITORII
Dup cum am vzut, transmisia impulsului nervos se
face cu ajutorul neurotransmittorilor. n SN sunt pro-
duse numeroase substante neurotransmittoare, fiecare
neuron elibernd una sau chiar mai multe. Au fost des-
coperiti n jur de 70 de neurotransmittori, a cror
actiune se cunoaste si pn la cca. 200 a cror actiune
nu se cunoaste nc. Unii neurotransmittori au efect
excitator pentru anumiti receptori si inhibitor pentru
altii, deoarece moleculele receptoare sunt diferite. De
regul, neurotransmittorii sunt mprtiti n dou grupe:
Curs de neuropsihologie 12
1) neurotransmittori cu molecul mic i actiune rapi-
d (tabelul 2.1.) si 2) neurotransmittori cu molecul
mare i actiune lent sau neuropeptide.
Acetilcolina (ACh) a fost primul neurotransmi-
ttor descoperit (n 1914, de ctre Henry Hallett Dale si
confirmat ca neurotransmittor de ctre Otto Loewi,
ambii cercettori primind Premiul Nobel pentru
Fiziologie sau Medicin, n 1936), unicul cu molecul
mic si actiune rapid care nu este nici aminoacid, nici
derivat de aminoacid. Este secretat din multe regiuni,
dar mai ales de 1) cortexul motor, 2) anumiti neuroni
din ganglionii bazali, 3) neuronii motori ai muschilor
scheletici, 4) unii neuroni din SNV. S-au identificat, n
creier, dou sisteme colinergice (care produc ACh)
majore : unul reticulat ascendent (continuare a forma-
tiunii reticulate din trunchiul cerebral) si altul limbic
(centrat pe hipocamp).
Actiunea fiziologic a ACh asupra muschilor
este de a-i activa, adic de a le provoca o contractie.
Actiunea central (din SNC) a activittii ACh se mani-
fest sub forma trezirii corticale si comportamentale si
asupra memoriei. De cele mai multe ori, ACh are un rol
excitator, dar poate avea si un rol inhibitor, de exemplu
inhibitia inimii realizat de nervii vagi (SNV parasim-
patic).
Anticorpii care blocheaz receptorii ACh
cauzeaz miastenia gravis, o boal caracterizat prin
oboseal si slbirea tonusului muscular. n trecut, pro-
voca moartea bolnavului, dar medicina modern are
metode pentru a controla boala, ns nu si pentru a o
vindeca. Alt boal n care este implicat ACh este boa-
la Alzheimer. Aceast dement se caracterizeaz prin
deficiente cognitive, n special mnezice, care coreleaz
cu scderea productiei de ACh din hipocamp. Cu ct
boala avanseaz, cu att hipocampul produce mai
putin ACh.
Norepinefrina este o monoamin, derivat al
dopaminei si este secretat, n SNC, de numerosi neu-
roni situati n trunchiul cerebral (n special la nivelul
puntii) si hipotalamus. Efectul psihologic al norepine-
frinei este legat de controlul de ansamblu al activittii si
al strii de dispozitie, realiznd, de exemplu, cresterea
nivelului de alert. n majoritatea ariilor cerebrale unde
actioneaz, norepinefrina are rol excitator, ns, n anu-
mite regiuni, poate activa si receptori inhibitori.
Cocaina si amfetaminele prelungesc actiunea norepine-
frinei, neuronii receptori rmnnd activati pentru pe-
rioade mai lungi de timp, ceea ce confer efectul psiho-
logic stimulativ al acestor droguri.
n SNP este implicat n realizarea sinapselor
pre- si postganglionare (SNV simpatic), stimulnd anu-
mite organe si inhibnd altele.
Epinefrina, hormon secretat de glanda supra-
renal si mediator simpatic, determin efecte similare
norepinefrinei, excitatorii, asupra SNV simpatic : vaso-
constrictie (ngustarea vaselor sanguine), stimularea ini-
mii, bronhodilatatie si unele efecte metabolice. Epine-
frina si norepinefrina sunt catecolamine.
Dopamina (DA) este o monoamin din clasa
catecolaminelor. Exist cel putin trei ci secretoare de
DA : 1) de la substanta neagr din trunchiul cerebral la
ganglionii bazali (cale responsabil si foarte afectat n
cazul bolii Parkinson ; bolnavii, practic, nu au DA n
creier); 2) din apropierea substantei negre pn la
bulbul olfactiv al sistemului limbic (foarte afectat n
cazul schizofreniei, medicamentele pentru aceast boal
blocnd descrcarea DA ; cale implicat n memorie si
cognitie) ; 3) n hipotalamus (neuroni implicati n
controlul secretiei glandei pituitare, hipofiza).
Serotonina (5-HT) este o monoamin care, ca
si norepinefrina, joac un rol important n reglarea dis-
pozitiei. Este secretat de nucleii rafeului median din
mezencefal si se proiecteaz asupra multor regiuni ale
SNC, n special la nivelul coarnelor posterioare ale
mduvei spinrii si n hipotalamus. Nivelurile sczute
de 5-HT sunt asociate cu depresia, antidepresivele
(Prozac, Zoloft, Paxil) nefiind altceva dect substante
care inhib reabsorbtia 5-HT. De asemenea, 5-HT are
un rol si n reglarea somnului, precum si n tratamentul
bulimiei. La nivelul mduvei spinrii, actioneaz ca un
inhibitor al cilor de transmitere a durerii.
Histamina se gseste n hipotalamus, dar si n
alti nuclei cerebrali. Are un rol important n diminuarea
reactiilor inflamatorii, controleaz permeabilitatea vase-
lor de snge si musculatura neted si are un rol impor-
tant n functionarea glandelor exocrine. Din histamin
deriv dou peptide.
Clasa Neurotransmitorul
Clasa I Acetilcolin (ACh)
Clasa a II-a
Monoamine
Norepinefrina (noradrenalina)
Epinefrina (adrenalina)
Dopamina (DA)
Serotonina (5-HT)
Histamina
Clasa a III-a
Aminoacizi
Acidul gamma-aminobutiric (GABA)
Glicina
Glutamatul
Clasa a IV-a Oxidul nitric (NO)
Tabelul 2.1. Neurotransmitori cu molecul mic i
aciune rapid
13 Curs de neuropsihologie
Aminoacizii sunt constituenti celulari universali
si sunt, prin urmare, produsi n toate celulele nervoase.
Nu se stiu foarte multe n legtur cu activitatea lor ca
neurotransmittori, ns au, cu sigurant, un rol modu-
lator asupra activittii altor neuroni. Glutamatul,
aspartatul, glicina si GABA sunt patru din cei 20 de
aminoacizi comuni tuturor celulelor. Glutamatul este
considerat principalul modulator excitator al sinapselor
SNC, iar GABA principalul inhibitor.
Glutamatul este probabil secretat de terminatiile
presinaptice din numeroase ci senzitive, precum si n
multe arii corticale, fiind un mediator excitator. Nu se
cunoaste precis rolul aspartatului, dar se presupune c
este tot unul excitator.
Glicina este secretat n principal la nivelul
sinapselor medulare si este posibil s actioneze mereu
camediator inhibitor.
Acidul gamma-aminobutiric (GABA) este sin-
tetizat din glutamat si este prezent n concentratie foarte
mare la nivelul SNC, dar poate fi depistat si n alte tesu-
turi. Principalul modulator inhibitor este secretat de
mduva spinrii, cerebel, ganglionii bazali si alte zone
corticale. n maladia Huntington, un sindrom ereditar
care se activeaz pe la 40 de ani, se distrug neuronii
GABA-ergici care coordoneaz miscarea, cauznd, prin
urmare, miscri necontrolate.
Oxidul nitric (NO)
1
este un gaz, secretat n
special de terminatiile nervoase din zone ale creierului
responsabile pentru comportamentul pe termen lung si
pentru memorie. NO nu este preformat si stocat n
veziculele din butonii terminali presinaptici, cum este
cazul altor neurotransmittori, ci este sintetizat aproape
instantaneu cnd este necesar si difuzeaz spre exterio-
rul terminatiilor presinaptice pe parcursul unui interval
de cteva secunde. La nivelul neuronului postsinaptic,
nu altereaz potentialul de membran, ci modific, pen-
tru un interval de cteva secunde sau minute functiile
metabolice intracelulare care influenteaz excitabilita-
tea neuronal. Medicamentul Viagra (sildenafil citrat)
este folosit, pe scar larg, pentru disfunctiile erectile
masculine si actioneaz prin mbunttirea activittii
NO.
Peptidele reprezint o clas complet diferit de
neurotransmittori, care sunt sintetizati diferit si ale c-
ror actiuni sunt, de obicei, lente si, n anumte privinte,
semnificativ diferite de actiunile neurotransmittorilor
cu molecul mic. Spre deosebire de substantele din
prima categorie, care sunt sintetizate, de regul, n
butonii terminali, peptidele iau nastere la nivelul reti-
1
Nepredat la curs.
culului endoplasmatic (din corpul celular), de unde se
ndreapt ctre aparatul Golgi, unde are loc definitiva-
rea procesului de formare. Ulterior, sub form de granu-
le secretorii, prsesc aparatul Golgi, ndreptndu-se
lent, cu o rat de numai ctiva cm/zi, prin transport
axonal, ctre terminatiile axonale (butonii terminali). n
neuroni, au fost descoperite mai mult de 50 de peptide.
Cnd actioneaz la distant, peptidele se
numesc hormoni, iar cnd actioneaz local se numesc
neurotransmittori.
Studierea peptidelor este important si pentru
faptul c acestea sunt implicate n mecanismele sensibi-
littii si emotiilor. Exist mai multe familii de peptide,
unele din cele mai importantae fiind prezentate n
tabelul 2.2.
Tabelul 2.2. Cele mai importante familii de neuropeptide.
Din punct de vedere al transmiterii impulsului
nervos, peptidele pot avea att rol excitator, ct si rol
inhibitor. Peptidele pot actiona ca neurotransmittori (n
regiunile cerebrale implicate n perceptia durerii), ca
modulatoare si au rol n reglarea rspunsurilor la stress.
Opioidele actioneaz ca si opium-ul pentru a n-
ltura durerea sau cauza somnolent. n 1975, cercet-
torii au descoperit c creierul secret o substant asem-
ntoare ca efect cu morfina (enkefalin). Apoi au fost
descoperite alte opioide numite endorfine, care sunt de
trei feluri : u, si . u- si -endorfinele (se gsesc n
glanda pituitar) sunt similare cu enkefalina, iar -
endorfinele sunt mult mai puternice dect morfina si, pe
deasupra, reprezint un factor de eliberare pentru
hormonul de crestere si prolactin. Putinele cercetri au
subliniat faptul c endorfinele ajut la mentinerea
comportamentului normal. Endorfinele au un rol impor-
tant si n controlul secretiei de insulin, glaucagon,
hormoni tiroidieni si suprarenali. De asemenea, ele
intervin si n functia glandelor sexuale.
Familia Peptidele
Opioide Opicortine, Enkefaline, Dinorfina
Neurohipofizare Vasopresina, Oxitocina, Neurofizinele
Secretine Secretina, Glaucagonul, Peptida
vasoactiv intestinal, Peptida
inhibitoare gastric, Factorul de
eliberare a hormonului de cretere
(GHrH), Peptida izoleucinamida
histidinic
Insuline Insulina, Insulina ca factor de cretere I
i II
Somatostatine Somatostatinul, Polipeptida pancreatic
Gastrine Gastrina, Colecistochinina
14 Curs de neuropsihologie
3. ANATOMIA FUNCIONAL A SISTEMULUI NERVOS
CENTRAL UMAN
Introducere
1
. Pentru a ntelege comportamen-
tul uman este necesar s investigm modul de organiza-
re functional si anatomic a sistemului nervos (SN).
Acesta dispune de o arhitectur complex, guvernat de
seturi relativ simple de principii functionale, organiza-
tionale si de dezvoltare.
Criteriile de delimitare a segmentelor si forma-
tiunilor care care alctuiesc SN sunt diverse. Cea mai
simpl delimitare (topografic) mparte SN n dou
componente : sistemul nervos central (SNC) si sistemul
nervos periferic (SNP). SNC se compune din creier si
mduva spinrii, iar SNP din sistemul nervos somatic,
care transport mesaje la si de la receptorii organelor de
simt, muschi si suprafata corpului si sistemul nervos
vegetativ sau autonom (SNV/SNA), care asigur
coordonarea functionrii organelor interne si glandelor.
La rndul su, SNV se mparte n sistemul nervos
simpatic si sistemul nervos parasimpatic (fig. 3.1).
SNC al adultului poate fi mprtit n cinci divi-
ziuni : 1) mduva spinrii, 2) trunchiul cerebral, 3)
cerebelul, 4) diencefalul si 5) telencefalul (fig. 3.1).
SNC este sustinut si protejat de ctremeninge,
trei membrane conjunctive situate ntre creier si oasele
1
Facem precizarea c aceast introducere nu a fost predat la
curs, deci studentul poate trece peste ea, ns considerm c
este esential pentru ntelegerea unittilor de curs ce urmeaz
si, n general, pentru ntelegerea anatomiei sistemului nervos.
craniene si ntre mduva spinrii si coloana vertebral.
La exterior se situeaz dura mater (o membran fibroa-
s puternic, cu dou straturi), la mijloc arahnoida (o
membran subtire, delicat ce nconjoar SNC), iar la
interior pia mater (o membran subtire ce nveleste
intim SNC, este bogat vascularizat si contine vase mici
ce irig SNC). Spatiul situat ntre pia mater si arahnoid
se numeste spatiul subarahnoid si este umplut cu un
lichid numit lichidul cerebrospinal (LCS) (fig. 3.2).
SNP este acoperit cu dou meninge. Arahnoida,
mpreun cu santul ei asociat de LCS, este prezent
doar n SNC. n afara SNC, meningele exterior (dura
mater) si cel inferior (pia mater) se unesc si formeaz o
Sistemul nervos
Sistemul nervos
central
Creier
Trunchiul cerebral
Cerebelul
Diencefalul
Telencefalul
duva spinrii
Sistemul nervos
periferic
Sistemul nervos
somatic
Sistemul nervos
vegetativ sau
autonom
Sistemul nervos simpatic
Sistemul nervos parasimpatic
Fig. 3.1. Schema organizrii sistemului nervos.
Fig. 3.2. Meningele.
Curs de neuropsihologie 15
teac ce acoper nervii cranieni si spinali, precum si
ganglionii periferici.
Pentru a descrie o structur anatomic att de
complex precum este creierul, trebuie s folosim ter-
meni care s indice directiile spatiale. Directiile din SN
sunt, n mod normal, descrise n functie denevrax, o li-
nie imaginar trasat de la mduva spinrii spre partea
frontal a creierului. Termenii de orientare spatial si
directiile pe care le indic se pot vedea n fig. 3.3.
Pentru a descoperi ce se afl n interiorul SN,
acesta a trebuit s fie tiat. Sectionarea SN nu se face la
ntmplare, ci n anumite moduri standard. Astfel, vom
avea, n plan orizontal sectiunea transversal, iar n
plan vertical sectiunea sagital saumediosagital (de
exemplu, diviziunea emisferelor). Sectiunea frontal
desparte partea frontal a creierului pentru a vizualiza
structurile interne (fig. 3.4).
Creierul este, de regul, mprtit n trei regiuni:
1) rombencefal sau creierul posterior (toate structurile
aflate n partea posterioar a creierului, ct mai aproape
de mduv), 2) mezencefal sau creierul mijlociu (partea
de mijloc a creierului) si 3) prozenecefal sau creierul
anterior (structurile localizate n partea anterioar a
creierului).
Creierul este foarte fragil si moale, iar conside-
rabila lui greutate (aprox. 1.400 gr.), la care se adaug o
constructie delicat, cere ca acesta s fie protejat de
socuri. De fapt, creierul uman nici nu-si poate suporta
propria greutate, motiv pentru care, atunci cnd este
scos din cutia cranian, se turteste. Din fericire, cnd
este la locul lui, este bine protejat, datorit LCS din
spatiul subarahnoid, n care acesta pluteste. Fiind
scufundat n lichid, greutatea creierului este amortizat
semnificativ (ajunge pn la 80 gr. !), reducnd astfel
presiunea asupra bazei. LCS reduce, de asemenea, si
socurile pe care le-ar resimti SNC la ntoarcerea rapid
a capului. LCS este extras din snge si are o compozitie
asemntoare cu cea a plasmei sanguine. Este produs de
ctre un tesut special, cu o bogat irigare sanguin, nu-
mit plexul coroid, situat n cel de-al treilea ventricul.
Ventriculii cerebrali sunt niste scobituri,
niste camere interconectate, pline cu LCS. Cele mai
mari sunt ventriculele laterale (ventriculul I si II, dar nu
se foloseste terminologia aceasta), care sunt conectate
cu cel de-al III-lea ventricul (V3). Peretii V3 divid
partea nconjurtoare a creierului n jumtti simetrice.
Massa intermedia (adeziunea intertalamic vezi cap.
3.4.1.), o mas de tesut nervos care uneste cei corpi
talamici, strbate mijlocul V3 si este un convenabil
punct de referint. Ventriculul IV este unit cu V3 prin
apeductul cerebral sauapeductul lui Sylvius (fig. 3.5).
Fig. 3.3. Direciile de orientare spaial a SNC.
Fig. 3.5. Ventriculii cerebrali.
Fig. 3.4. Secionarea standard a SNC.
Curs de neuropsihologie 16
3.1. MDUVA SPINRII
Mduva spinrii este componenta SNC cea mai de jos
(distal inferioar), este situat n coloana vertebral si
este formatiunea cea mai veche filogenetic. Coloana
vertebral prezint 33 sau 34 de vertebre, mprtite n
cinci regiuni : 7 vertebre cervicale, 12 toracice, 5
lombare si 4 sau 5 coccigiene. Mduva spinrii are
forma unui cilindru usor turtit si se ntinde pe o lungime
de aproximativ 43-45 cm, de la vertebra Atlas (C1)
pn la a doua vertebr lombar (L2), terminndu-se
printr-un con conul medular (fig. 3.6). n partea
superioar, mduva spinrii se continu cu trunchiul
cerebral si este nconjurat de cele trei meninge (vezi p.
14), iar arahnoida este locul n care se produc cele mai
multe tumori. n partea de jos, dura mater formeaz
sacul dural, locul unde se fac punctiile lombare.
Mduva spinrii este compus din 31 de segmente, iar
fiecare segment d nastere la cte o pereche de nervi
spinali, care conecteaz diferite prti ale corpului cu
SNC (deci fac parte din SNP).
Structura intern a mduvei spinrii relev,
printr-o sectiune transversal (fig. 3.4), substanta
cenuie n interior si substanta alb, la exterior,
mprejurul celei cenusii
1
. Aceast dispunere nu este una
ntmpltoare, deoarece functiile principale ale mduvei
spinrii sunt de a trimite fibre motorii ctre efectori
(muschi si glande) si de a capta si transmite ctre creier
informatiile venite pe calea somatosenzorial (de la
receptorii senzoriali). Prin urmare, pentru realizarea
acestor functii, de captare si transmitere a semnalelor,
este normal s existe substant alb la exterior (pentru
captare si transmisie) si substant cenusie la interior
(pentru prelucrarea informatiei).
O alt particularitate a structurii anatomice
interne a mduvei este dispunereasubstanei cenuii n
form de H sau de fluture (fig. 3.7). Aceasta o face s
fie divizat n patru mari regiuni : coarnele posterioare
(dorsale), coarnele anterioare (ventrale), zonele
intermediare si coarnele laterale
2
.
Coarnele posterioare contin grupuri de neuroni
ce primesc impulsuri senzoriale de la rdcinile
1
n SN, substanta (materia) cenusie este alctuit dintr-un
conglomerat de milioane de corpuri celulare neuronale, iar
substanta alb din milioane de axoni. Putem compara
substanta cenusie cu un calculator si substanta alb cu firele
care ies din el, din punct de vedere functional.
2
n literatura de specialitate, se mai gsesc cu numele de
coloane cenuii, iar cordoanele din substanta alb (vezi
pagina urm.), se mai ntlnesc cu nunele decoloane albe.
posterioare (ale nervilor), ceea ce nseamn c coarnele
posterioare reprezint partea senzorial a substantei
cenusii spinale.
Coarnele anterioare contin grupuri de neuroni
ce au rol n miscrile voluntare, iar axonii lor prsesc
mduva prinrdcinile anterioare.
Zonele intermediare sunt situate ntre coarnele
anterioare si cele posterioare si sunt alctuite din
neuroni de asociatie, motiv pentru care zona
intermediar devine partea de asociatie a mduvei.
Majoritatea axonilor acestor neuroni nu prsesc
mduva, iar ctiva trimit proiectii spre creier.
Coarnele laterale sunt niste mici extensii, de
form triunghiular, ale zonelor intermediare. Contin
corpi neuronali ai preganglionilor din sistemul nervos
simpatic (SNV).
Neuronii substantei cenutii sunt aseazati
stratificat, cptnd un aspect laminar. Laminele
furnizeaz o identificare mai precis a ariilor substantei
cenusii si sunt foarte folositoare n descrierea locatiilor
originilor sau terminatiilor cilor functionale. Exist
zece lamine, numerotate dinspre posterior ctre
anterior, astfel : conul posterior contine laminele I-IV,
zona intermediar este, n
Fig. 3.6. ezarea mduvei spinrii n coloana vertebral.
Curs de neuropsihologie 17
principal, lamina VII, iar conul anterior contine o parte
din lamina VII si laminele VIII-X. Lamina X este aria
comisural din jurul canalului central.
Substana alb, nconjurnd substanta cenusie,
este divizat n trei regiuni, denumitecordoane. La fel
ca si coarnele substantei cenusii, cordoanele pot fi
anterioare, posterioare si laterale.
Cordoanele anterioare sunt situate ntre antul
median anterior si coarnele anterioare si contin fibre
nervoase (fascicule de axoni) care conduc impulsuri
motorii.
Cordoanele posterioare sunt situate ntre santul
posteromedial si coarnele posterioare. Ele dispun de
dou tipuri de tracturi (mnunchiuri de nervi) :
ascendente (Goll si Burdach) si descendente. Transport
impulsuri senzoriale.
Cordoanele laterale, situate ntre coarnele
anterioare si cele posterioare, contin tracturi des-
cendente (corticospinal/piramidal ncrucisat, rubro-
spinal, olivospinal, vestibulospinal, reticulospinal
lateral), tracturi ascendente (spinocerebelos dorsal,
spinocerebelos ventral, spinotalamic lateral, spino-
tectal) si tracturi de asociatie (intersegmentar lateral).
Se ocup de functionarea SNV simpatic.
Nervii spinali se ataseaz (n afar de primul si
ultimul) de mduva spinrii prin rdcinile anterioare
(ventrale) si cele posterioare (dorsale). Rdcina
anterioar si are originea real n neuronii motori (care
fac sinaps cu muschii scheletici sau netezi prin axonii
din nerv) din cornul anterior, iar rdcina posterioar n
ganglionul spinal. Rdcinile anterioare si cele
posterioare se unesc imediat dup ganglionul spinal
(fig. 3.7), formnd nervii spinali, care apoi se ramific,
lund calea diferitelor zone din corp.
Exist dou mari i senzoriale ce trec prin
mduva spinrii si ajung la creier : 1) sensibilitatea
cutanat (receptorii senzoriali din piele transmit, prin
nervi, semnalele de atingere, termice, de durere algice
la emisfera contralateral
1
) si 2) sensibilitatea
proprioceptiv (semnalele ajung la emisfera cerebral
contralateral si la emisfera cerebeloas ipsilateral
2
).
ile motorii transmit impulsuri de la creier sau
chiar din mduv (situatie n care avem de-a face cu
reflexe) ctre muschi sau organele efectoare. Miscrile
provocate pot fi voluntare (semnalele provin din
structurile superioare ale creierului) sau involuntare (de
la structurile subcorticale sau mduva spinrii).
Tracturile motorii de la creier la muschi sunt de dou
feluri : 1) tractul piramidal (corticospinal) sau 2) extra-
piramidal (tracturi provenite de la alte structuri
cerebrale, subcorticale, cum ar fi nucleii bazali sau
talamusul). Miscrile involuntare tin de activitatea
muscular n legtur cu pozitia corpului si echilibrul,
coordonarea miscrilor si a tonusului muscular.
La nivelul mduvei se poate constata si o
anumit autonomie motorie (adic miscri care nu sunt
provocate de structurile cerebrale) care const n
reflexele spinale. Acestea se compun din trei elemente :
1) neuroniii senzitivi, 2) neuronii asociativi din mduva
spinrii si 3) neuronii motori. Un act reflex este un
rspuns imediat la un stimul. Informatia senzorial
ptrunde prin coarnele posterioare, de unde este
preluat de neuronii asociativi si transmis neuronilor
motori din coarnele anterioare, care provoac o miscare.
De exemplu, cnd ne este testat reflexul rotulian (cu
1
Situat de cealalt parte. De exemplu, senzatiile de la mna
stng ajung n emisfera dreapt.
2
De aceeasi parte.
Fig. 3.7. Structura intern a mduvei spinrii i formarea nervilor spinali.
Curs de neuropsihologie 18
ciocnelul n genunchi), informatia senzorial
provocat de lovitura ciocnelului cltoreste prin
intermediul cii senzitive (aferente) a nervului spre
partea noastr din spate, unde intr n mduva spinrii
si provoac un rspuns imediat, ce va veni pe calea
motorie (eferent) la doi muschi care vor mpinge
partea de jos a piciorului nainte si vor retrage
genunchiul (fig. 4.8).
Patologia mduvei spinrii
Leziunile nervilor periferici (spinali) dau nastere la
tulburri motorii, tulburri senzoriale sau ambele.
Aceste leziuni se grupeaz sub numele de neuropatie
periferic. Aceasta poate fi :
- Radiculopatia (lat. radix rdcin) este rezultatul
lezrii unei rdcini nervoase. Cea mai rspndit
cauz estespondiloza, care provoac lezarea uneia
sau mai multor rdcini nervoase. Lezarea unei
rdcini dorsale produce abolirea impulsurilor
senzoriale care vin ctre mduv si, implict,
ntreruperea buclei reflexelor spinale. Lezarea unor
rdcini ventrale produce deficite n rspunsul
motor.
- Mononeuropatia reprezint deficitul ce reflect
lezarea unui singur nerv periferic. Cele mai
frecvente cauze sunt traumatismele (loviturile
fizice puternice).
- Polineuropatia include deficite senzoriale si
motorii care reflect lezarea mai multor nervi
periferici.
Neuropatia periferic survine deseori la diabetici sau la
persoanele care sufer de boli autoimune precum artrita
reumatoid sau lupus. Unele deficiente de vitamine,
medicamente si alcoolismul pot, de asemenea, s
afecteze nervii periferici.
Alte afectiuni ale mduvei spinrii :
- Tumorile subdurale sunt excrescente ce apar sub
dura mater.
- Tumorile medulare apar n interiorul substantei
cenusii sau albe si sunt, de cele mai multe ori,
metastaze.
- Aparitia unor tumori vasculare (pe vasele de
snge).
- Inflamatia rdcinilor nervilor spinali provoac
diverste tipuri de dureri nevralgice sau mialgice
(reumatismul) legate de pierderea sensibilittii. De
asemenea, bolnavul poate acuza probleme de tonus
muscular si pot aprea paraliziile partiale (para-
parez) sau care implic muschii tuturor mem-
brelor (tetraparez).
Eforturile psihologice de recuperare sunt
necesare si enorme. Pot aprea nevroze, forme usoare
sau grave de depresie si/sau confuzie, apatie si
agresivitate, extra- si interpunitivitate. Se fac terapii n
grup si sedinte de relaxare.
3.2. TRUNCHIUL CEREBRAL
Creierul este cea de-a doua mare component a SNC.
Acesta se afl n prelungirea mduvei spinrii si este
situat n cutia cranian, protejat de cele trei meninge
(fig. 3.2). Exist diferite moduri de a diviza creierul n
substructuri anatomice, dup cum ziceam la nceputul
acestui capitol, iar modul ales de noi l mparte n :
trunchiul cerebral, cerebel, diencefal si telencefal,
fiecare structur avnd substructurile ei, unele din ele
putnd fi vizualizate n fig. 3.9.
Trunchiul cerebral este partea cea mai de jos a
creierului, aflndu-se n continuarea mduvei spinrii si
este acoperit, n partea posterioar, de ctre cerebel, cu
care este conectat prin mase extrem de mari de fibre
nervoase ce formeaz trei perechi depedunculi cerebe-
loi. n partea superioar a trunchiului cerebral se afl
diencefalul, de care este desprtit printr-o comisur
(zon plin de fibre nervoase, prin intermediul creia
comunic emisferele cerebrale).
Fig. 3.8. Mecanismul de funcionare a reflexului spinal.
Curs de neuropsihologie 19
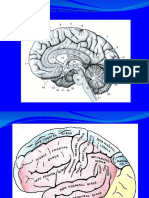
Fig. 3.9. Seciune mediosagital prin creier, evideniind principalele componente anatomice ale acestuia.
Fig. 3.10. Seciune anterioa prin creier ce relev structura exterioar anterioar a trunchiului cerebral i a unor
componente aflate n apropierea acestuia.
Curs de neuropsihologie 20
Structura intern a trunchiului cerebral este ase-
mntoare cu cea a mduvei spinrii, adic prezint
substant cenusie la interior, nconjurat de substant
alb.
Pe suprafata trunchiului cerebral se gseste
originea aparent
1
a ultimelor zece perechi de nervi
cranieni (III-XII) (fig. 3.10).
Trunchiul cerebral este alctuit din componente
supraetajate. De jos n sus acestea sunt : bulbul rahi-
dian (sau medulla oblongata), pons (protuberanta,
puntea lui Varolio sau pur si simplu puntea) si
mezecncefalul (fig. 3.9). De asemnea, n centrul
trunchiului cerebral, se afl formatiunea sau subtanta
reticulat, care se ntinde de la bulbul rahidian pn la
partea superioar a mezencefalului.
3.2.1. Bulbul rahidian (medulla oblongata)
Termenul de medulla oblongata ar nsemna,
literal, un element central alungit. Dup cum se poate
vedea n fig. 3.9 - 3.11, medulla oblongata are, ntr-
adevr, o form alungit si se afl ntr-o pozitie central
a nevraxului (p.15, stnga sus detalii despre nevrax).
Bulbul rahidian (BR) este partea cea mai de jos
a trunchiului cerebral, situndu-se deasupra si n prelun-
girea mduvei spinrii. n partea superioar, ajunge
pn la punte, de care este desprtit prinantul bulbo-
pontin (un sant transversal). Are forma unui trunchi de
con, cu baza mic (diametru 1,25 cm) nspre mduv
si baza mare (d 2 cm) nspre punte. Lungimea BR este
de aproximativ 3 cm.
La exterior, bulbul prezint patru fete: una
anterioar, una posterioar si dou laterale.
Pefata anterioar (fig. 3.10), de o parte si de
alta a antului median anterior, se gsesc piramidele
bulbare, prti componente ale tractului piramidal
(corticospinal), prin care circul impulsuri nervoase
descendente (motorii). Neuronii motori, care pleac din
zonele superioare, se ncruciseaz la nivelul bulbului,
trecnd dintr-o parte n cealalt (n proportie de 80%),
formnd astfel decusatia piramidal. Acest lucru face
ca emisfera dreapta s controleze miscrile din partea
stng a corpului, iar emisfera stng pe cele din partea
dreapt. Imediat deasupra piramidelor bulbare, n santul
bulbopontin, si are originea aparent nervul abducens
(VI).
De asemenea, si unele tracturi ascendente (sen-
zoriale) sedecuseaz la nivelul bulbului, iar altele, mai
jos, n mduva spinrii.
1
Originea aparent difer de cea real, prima fiind portiunea
exterioar, vizibil din care iese nervul, iar cea real este
reprezentat de nucleii aflati n interior.
Pe fetele laterale se gsesc rdcinile nervului
glosofaringean (IX) si vag (X), precum si olivele bul-
bare, care contin un nucleu de substant cenusie. ntre
piramidele si olivele bulbare se gseste originea aparen-
t a nervului hipoglos (XII). Deasupra olivei bulbare se
gseste emergenta aparent a nervului facial (VII), iar
deasupra emergentei nervului glosofaringean, se afl
emergenta aparent a nervului vestibulocohlear (VIII),
dup cum se poate vedea n fig. 3.10.
Fata posterioar (fig. 3.11), care nu este
vizibil, fiind acoperit de cerebel, prezint o structur
diferit n portiunea superioar fat de cea inferioar. n
partea inferioar, structura este asemntoare cu cea a
cordoanelor posterioare din mduva spinrii. n partea
superioar, fasciculele Goll si Burdach (tracturi ascen-
dente, senzoriale) se separ, tractul Burdach prelungin-
du-se cu pedunculul cerebelos inferior, iar tractul Goll
devenind piramida bulbar posterioar. n partea
superioar (a fetei posterioare) se gseste planseul
ventriculului IV. Aici se gsesc nucleii de origine ai
nervilor: glosofaringean (IX), vag (X), accesoriu (XI) si
hipoglos (XII).
Structura intern a bulbului este asemntoare
cu cea a mduvei spinrii din mai multe puncte de
vedere, n principal din cel al dispunerii substantelor
nervoase cenusie la interior si alb la exterior,
mprejurul celei cenusii. ns decusatia piramidal din
bulb si decusatia unor ci ascendente (senzitive) fac ca
substana cenuie s fiefragmentat, spre deosebire de
cea din mduva spinrii, dnd nastere unor mici insule
sau nuclei (conglomerate de corpi neuronali). Prin
fragmentarea coloanelor posterioare (vezi structura
intern a mduvei spinrii) se formeaz nucleii senzi-
tivi, prin fragmentarea coloanelor anterioare se formez
nucleii motori, iar prin fragmentarea coloanelor laterale
nucleii vegetativi. Nucleii formati prin segmentare se
numesc echivalenti (celor din mduv). n afar de
acestia, bulbul are si nuclei proprii.
Nucleii senzitivi sunt : nucleul vag (X), nucleul
glosofaringean (IX), o parte din nucleul trigemenului
(V), nucleul vestibulocohlear (VIII), nucleul solitar.
Nucleii motori sunt : nucleul nervului hipoglos
(XII) si nucleu ambiguu al nervilor glosofaringean
(IX), vag (X) si accesoriu (XI).
Nucleii vegetativi sunt : nucleul solitar inferior
(trimite fibre eferente ctre glandele salivare prin inter-
mediul nervului XII) si nucleul dorsal al nervului vag
(trimite fibre eferente la inim, plmni si organe abdo-
minale).
Curs de neuropsihologie 21
Nucleii proprii sunt cei n care sfrsesc tracturi-
le Goll si Burdach, fiind, prin urmare, implicati n pro-
cesarea sensibilittii proprioceptive.
Oliva bulbar este un nucleu ce are conexiuni
importante cu : cerebelul, nucleul rosu(din mezencefal),
corpii striati, creierul mare si mduva spinrii. Este un
nucleu care face parte din sistemul extrapiramidal si
particip la realizarea miscrilor involuntare.
Formatiunea reticulat este o structur alctuit
din multi nuclei (peste 90), ai cror neuroni sunt difuzi
si interconectati, formnd o retea complex. Ea ocup
centrul ntregului trunchi cerebral, pornind la baza BR
si ajungnd pn la limita superioar a mezencefalului.
Formatiunea reticulat primeste numeroase informatii
senzoriale si proiecteaz axoni ctre scoarta cerebral,
talamus si mduva spinrii. J oac un rol important pen-
tru somn, trezire (reactia de orientare a atentiei), tonus
muscular, miscare si diverse reflexe vitale. Va fi tratat
mai pe larg ntr-o unitate viitoare.
Substana alb a bulbului este format din fibre
mielinizate, grupate, dup cum am vzut, sub form de
tracturi. Tracturile bulbare sunt de dou feluri : 1) de
trecere (asecendente si descendente) si de asociatie si
2) proprii.
Tracturile ascendente, ce provin din mduv,
sunt : Goll, Burdach, spinocerebelos si spinotalamic.
Tracturile descendente provin din structurile
anatomice situate desupra bulbului : tractul piramidal
(corticospinal), rubrospinal, tectospinal, olivospinal,
reticulospinal, vestibulospinal (prin formatiunea reticu-
lat).
Tracturile asociative se reunesc n bulb, punte
si mezencefal, formnd fasciculul longitudinal medial,
care este situat posterior lemniscului medial.
Fibrele proprii si au originea n bulb si sunt
descrise sub forma diferitelor grupri.
Bulbul rahidian este o structur anatomic ner-
voas ce reuneste o multitudine de nuclei si fascicule
nervoase, ceea ce indic faptul c BR are o mare impor-
tant n functionarea organismului. Principalele funcii
ale BR sunt : 1) functia reflex (de asociatie), 2) functia
de conducere nervoas (substanta alb) si 3) functia de
reglare a tonusului si dinamicii activittii psihice
Functia reflex este responsabil pentru
controlul unor centri vitali : centrul respirator (functie
parasimpatic), centrul cardiac (asigur relaxarea ini-
mii), centrul vasomotor (asigur dilatarea si constrictia
vaselor sanguine), centrul salivatiei, centrul deglutitiei
(nghitirii), centrul suptului, centri ai unor reactii de
aprare (strnutul, tusea, clipitul, voma) si centri de re-
glare a tonusului muscular (intensificarea acestuia).
Functia de conducere const n transmiterea :
1) ntre zonele receptoare si centrii superiori ai sensibi-
littii (aferenti) si 2) ntre centrii de comand (motori) si
organele efectoare corespunztoare. Desigur, 1) se rea-
lizeaz prin intermediul tracturilor ascendente si 2) prin
intermediul tracturilor descendente.
Functia de reglare a tonusului i dinamicii
activittii psihice se realizeaz, cu precdere, prin inter-
mediul formatiunii reticulate. Formatiunea reticulat
este implicat n coordonarea muschilor scheletici,
Fig. 3.11. Seciune postero-lateral prin creier ce relev structura posterioar a trunchiului ceerebral i a
altor componente aflate n apropierea acestuia.
Curs de neuropsihologie 22
coordonarea activittii organelor (SNV) si n atentia
selectiv.
3.2.2. Pons (puntea lui Varolio)
Pons este o formatiune nervoas care apare la
mamifere si este cel mai dezvoltat la om. Este situat
anterior de cerebel, deasupra bulbului rahidian si dede-
suptul mezencefalului. Lateral, se ntinde de la o
emisfer cerebeloas la alta (Fig. 3.9, 3.10, 3.11). n
partea superioar, pons este desprtit de mezencefal
prin antul pontopeduncular, iar n partea inferioar
este desprtit de bulb prinantul bulbopontin.
Ca si bulbul rahidian, pons prezint, la exterior,
o fat anterioar, una posterioar si dou laterale.
Pe mijlocul fetei anterioare, exist un sant
longitudinal numit antul bazilar. Pe marginile santului
apar dou umflturi, numite piramidele pontine,
prelungiri ale piramidelor bulbare.
Fata posterioar (fig. 3.11) este acoperit de
cerebel, are o form triunghiular si constituie jumta-
tea superioar a planseului ventriculului IV.
Fetele laterale se continu cu pedunculii cere-
belosi mijlocii. La limita dintre pons si pedunculii
cerebelosi mijlocii se afl emergenta aparent a
nervului trigemen (V) (fig. 3.10, 3.11).
Structura intern prezint aceeasi organizare
ca mduva spinrii si bulbul rahidian substanta cenu-
sie la interior si substanta alb la exterior, mprejurul
substantei cenusii. Substanta cenusie este fragmentat
n nuclei, iar substanta alb este predominant la acest
nivel al trunchiului cerebral.
Sectionnd transversal puntea (vezi fig. 3.4),
distingem dou prti : 1) piciorul (situat anterior) si 2)
calota (situat posterior). Cele dou prti sunt desprtite
de corpul trapezoid. Pe linia median, se oberv o
ncrucisare de fibre, numit rafeu, care mparte pons n
dou prti simetrice.
Piciorul contine substant alb si substant
cenusie. Substanta alb a piciorului contine dou tipuri
de fibre nervoase : 1) longitudinale, care pornesc din
scoarta cerebral si se termin n pons (corticopontine)
sau n mduva spinrii (corticospinale) ; 2) transversale,
care pornesc din punte si se pierd n pedunculii cerebe-
losi mijlocii. Substanta cenuie a piciorului contine
nucleii fibrelor nervoase transversale (din substanta al-
b a piciorului) si nuclei n care se termin unele fibre
longitudinale care vin de la scoarta cerebral (cortico-
pontine).
Calota (sau tegmentul) este format din sub-
stant alb, substant cenusie si substant (formatiune)
reticulat. Substanta alb a calotei cuprinde 1) fibre
ascendente medulare (Goll si Burdach) care se duc spre
cerebel (tractul spinocereelos), spre nucleii subcorticali
(tractul spinotalamic, fasciculul longitudinal medial,
lemniscul medial)si bulbare (corticopontinocerebeloase)
si 2) fibre descendente corticale (tractul piramidal),
subcorticale (rubrospinal, rubroolivar, tectospinal, reti-
culospinal) si, n parte, de la cerebel.
Corpul trapezoid este format din fibre transver-
se, constituind o parte a cilor de conducere auditive.
Fasciculul longitudinal medial contine fibre
ascendente si descendente. Acest fascicul este o cale
important de conducere a impulsurilor legate de
miscrile de orientare a ochilor, capului si gtului.
Lemniscul medial (panglica lui Reil) este
format din fibre ascendente, de la mduv si bulb spre
tectum (din mezencefal) si talamus. Este asezat central,
iar n partea sa lateral se gseste lemniscul lateral,
constituit din fibre acustice.
n pons, si au originea nervii : V (trigemen
motor si senzitiv), VI (abducens), VII (facial), VIII
(vestibulocohlear).
Pons ndeplineste dou funcii : 1) functie
reflex (reflexul lacrimal, salivar, masticator, corneran,
audiooculogir ntoarcerea capului dup sursa sonor
sau luminoas , sudoripar si sebaceu ale fetei si capu-
lui, controlul mimicii expresive, a fetei, miscarea latera-
l a globilor oculari, tonusul muscular) si 2) functie de
conducere (asigur circulatia informatiei extrase din
mediul extern si cel intern al organismului ctre centrii
subcorticali si corticali si a mesajelor de comand n
sens descendent ctre muschi si organele efectoare si
este ndeplinit prin intermediul fasciculelor nervoase).
3.2.3. Mezencefalul (creierul mijlociu)
Mezencefalul este cea de-a treia structur anato-
mic nervoas a trunchiului cerebral, situat deasupra
puntii, de care se desparte prinantul pontopeduncular,
si dedesuptul diencefalului, de care este desprtit printr-
un plan care uneste comisura (o zon foarte larg ce
contine fibre nervoase care transmit informatia ntre
emisfere) posterioar a creierului cu marginea poste-
rioar a corpilor mamilari. Mezencefalul este o struc-
tur anatomic ce nconjoar apeductul cerebral
(Sylvius). Are o fat anterioar, una posterioar si dou
laterale. Printr-o sectiune transversal, determinm trei
Curs de neuropsihologie 23
regiuni ale mezencefalului 1) tectum sau lama cvadri-
gemen, n partea posterioar, 2) tegmentum sau calota,
la mijloc si 3) crus cerebri sau picioarele pedunculilor
cerebrali, n partea anterioar (fig. 3.10). 2) si 3)
formeaz pedunculii cerebrali (lat. pedunculus
piciorus).
Fata anterioar este singura vizibil ; pe mijloc
se gsestefosa interpeduncular, n profunzimea creia
se gseste substanta perforat posterioar (fig. 3.10).
n prtile laterale, fosa interpeduncular este delimitat
de picioarele pedunculilor cerebrali (crus cerebri), care
sunt continuri ale tractului piramidal (vezi puntea si
bulbul).
Fata posterioar nu este vizibil, fiind acoperi-
t de partea superioar a cerebelului si de lobii occipi-
tali (fig. 3.9). Pe fata posterioar se gseste lama
cvadrigeminal (tectum mezencefalic) care contine
coliculii cvadrigemeni, dispusi sub form de dou pe-
rechi a cte doi coliculi superiori si inferiori (fig.
3.11).
Fetele laterale corespund tegmentului si picio-
rului peduncular. ntre picior si tegment se afl antul
lateral al mezencefalului.
1) Pedunculii cerebrali (tegmentul +picioarele
pedunculare) contin substant alb si cenusie(fig. 3.12).
Picioarele pedunculilor (crus cerebri) sunt
alctuite numai din substant alb, reprezentnd
fascicule de fibre nervoase care pornesc din scoarta
cerebral si ajung n pons si bulb (geniculate,
corticopontine) si n mduv (piramidale).
Tegmentumul (calota) este format din substant
alb si cenusie si este cuprins ntre tectum si crus
cerebri. ntre tegmentum si crus cerebri se afl
substanta neagr. n partea de jos, tegmentul mezence-
falic se continu cu tegmentul pontin, iar n partea
rostral, se pierde n diencefal. Substanta alb este alc-
tuit din fibre ce pleac din talamus si hipotalamus, din
mduva spinrii, pons si fibre care pornesc sau se
opresc din si n nucleii mezencefalului.
Lemniscul medial din pons se prelungeste n
substanta alb a tegmentului mezencefalic, posterior de
portiunea lateral a substantei negre (fig. 3.12). Unele
fibre ale sale se opresc n substanta neagr, iar altele
urc spre talamus. Lemniscul lateral contine fibre care
se opresc n coliculii cvadrigemeni, iar cea mai mare
parte ajung lacorpul geniculat medial al talamusului.
Fasciculul longitudinal medial reprezint o
important cale de legtur ntre nucleii trunchiului
cerebral. Este situat n partea ventral a substantei cenu-
sii centrale si are un rol foarte important n coordonarea
miscrilor ochilor, capului si trunchiului.
n substanta alb a tegmentului se mai gsesc :
tractul spinotalamic, cerebelotegmental, tectonuclear si
tectospinal, tractul central al tegmentului sau fasciculul
central al calotei.
Substanta cenuie a tegmentului este dispus, n
parte, n jurul apeductului cerebral (Sylvius), motiv
pentru care se numeste periapeductal, si n nuclei
specifici mezencefalului. Printre substanta alb si sub-
stanta cenusie se gseste formatiunea reticulat.
Substana neagr este cel mai mare nucleu al
mezencefalului, are o structur compact si form de
semilun. Substanta neagr primeste aferente (impul-
suri senzoriale) de la toat suprafata corpului si de la
organele vizuale, auditive si olfactive. Eferentele sunt
trimise att ctre structurile corticale, ct si ctre cele
subcorticale. Din conexiunile pe care le stabileste,
rezult c substanta neagr are un rol important n inte-
grarea senzorial si n reglarea miscrilor fine.
Nucleul rou (Stilling) este situat deasupra
substantei negre, are o form oval si este rosu datorit
mpregnrii corpilor celulari cu oxizi de fier si prezentei
numeroaselor vase mici de snge. Are conexiuni att cu
formatiunile superioare, ct si cu cele inferioare. Este o
component important a tracturilor extrapiramidale,
avnd rolul de a inhiba centri bulbari si medulari, ns
activitatea lui este subordonat, la rndul ei, controlului
centrilor extrapiramdali superiori si scoartei cerebrale.
Att nucleul rosu, ct si substanta neagr controleaz
tonusul muscular.
n substanta cenusie a tegmentului se mai g-
sesc : nucleul trohlearului (IV), nucleul oculomotorului
(III), nucleu tractului mezencefalic al trigemenului (V),
nucleul interstitial (centrul subcortical al miscrilor
verticale si rotatorii ale ochilor), nucleul lui
Darkschewitsch, nucleul intercrural, nucleul dorsal al
tegmentului.
Fig. 3.12. Seciune transversal a mezencefalului la
nivelul coliculilor superiori.
Curs de neuropsihologie 24
2) Tectumul mezencefalic este format din lama
cvadrigemen cu doi coliculi superiori si doi inferiori
(cvadrigemeni), dup cum se poate vedea n fig. 3.11.
Numele de cvadrigemeni (patru gemeni) se datoreaz
structurii asemntoare a celor patru coliculi. Tectumul
are, n compozitia sa, att substant alb, ct si
substant cenusie.
Coliculii cvadrigemeni superiori au, n struc-
tura lor, substant alb si substant cenusie dispuse
alternativ. Se conecteaz cucorpii geniculati laterali (ai
talamusului), de la care primesc aferente optice (tractul
geniculotectal) si de la lobul occipital (corticotectal).
De asemenea, primesc aferente si de la mduv
(spinotectal). Trimit eferente ctre nucleii nervilor
cranieni din trunchiul cerebral (III, VII, XI) si ctre
mduva spinrii. Au rolul de a regla automat miscrile
oculare si a celor implicate n orientarea si concetrarea
vizual.
Coliculii cvadrigemeni inferiori se situeaz sub
cei superiori, au dimensiuni ceva mai reduse si form
oval. Contin substant cenusie la interior, si substant
alb la exterior. Primesc aferente de la mduva spinrii
si de la lemniscul lateral (auditive). Trimit eferente
ctre corpii geniculati mediali (din talamus), fiecare
colicul trimite eferente ctre cel opus si ctre cel
superior, ctre pons, bulb si mduv. Au rol n realiza-
rea reflexelor de orientare la stimuli auditivi, reflex care
preced si faciliteaz perceptia auditiv.
Per ansamblu, mezencefalul ndeplinestefuncii
legate de reflexele de orientare, reflexele de redresare si
distributia normal a tonusului muscular.
3.2.4. Formaiunea reticulat (FR)
Printre tracturile si nucleii ce se afl n interio-
rul trunchiului cerebral, se afl o retea complex de
neroni ce poart numele de formatiune sau substant
reticulat.
FR se gseste la toate cele trei niveluri ale trun-
chiului cerebral, urcnd, n partea superioar, pn la
diencefal si cortex, si cobornd, n partea inferioar,
pn la mduva spinrii. A fost descoperit n 1911 de
ctre Santiago Ramn y Cajal (1852-1834), laureat al
Premiului Nobel pentru Medicin sau Fiziologie si
considerat unul dintre fondatorii neurostiintelor. Cnd a
fost descoperit, FR a fost descris ca simpl substant
de sprijin. De atunci, studiile au artat c este implicat
n multe alte functii, c are proiectii ctre centrii
superiori (F. Bremer, 1935), iar n 1954 i-au fost des-
crisi (Olszewski) 98 de nuclei a cror functie nu este
cunoscut. n 1965, Sager si colaboratorii si deosebesc
cinci grupe de nuclei : 1) nuclei care au legturi numai
cu cerebelul, 2) nuclei mediali cu axoni lungi ascenden-
ti si descendenti, 3) nuclei laterali cu rol de receptie si
asociatie, 4) nucleii rafeului cu rol n tonusul muscular,
5) nucleii lui Nauta. Se pare c un axon apartinnd FR
poate intra n contact cu 27.500 de alti neuroni! Aceast
capacitate face din FR o substant cu rol fundamental
n transmiterea i difuzarea rapid a informatiilor.
Asezarea central a FR n interiorul trunchiului
cerebral (fig. 3.13), sugereaz asocierea acesteia cu
cile ascendente, descendente si cu nucleii nervilor
cranieni. ntr-adevr, FR primeste aferente din toate
prtile SN si exercit influente aproape asupra fiecrei
functii a SNC.
n ziua de azi, se vorbeste de dou tipuri de FR:
1) formatiunea reticulat ascendent si 2) formatiunea
reticulat descendent, fiecare din ele avnd o compo-
nent activatoare si una inhibitoare. Astfel, se formeaz
patru sisteme : sistemul reticulat activator/inhibitor
ascendent (SRAA/SRIA) si sistemul reticulat
activator/inhibitor descendent (SRAD/SRID).
SRAA a fost descoperit de Moruzzi si Magoun
(1949), care au demonstrat c animalele adormite pot fi
trezite prin excitarea FR de la nivelul trunchiului cere-
bral si de la nivelul diencefalului. Prin urmare, SRAA
se ntinde de la trunchiul cerebral pn la scoarta cere-
bral si are o activitatetonic (mentine starea de veghe)
si o activitatefazic (activarea difuz a scoartei la actiu-
nea unui stimul). Acestea se mai gsesc, n literatura, de
specialitate cu numele de activare de fond (tonic) si
activare indus sau provocat (fazic). Orice disfunctie
a SRAA (n sensul scderii activittii) duce la somno-
lent permanent, iar functionarea normal a SRAA
este asociat cu starea de veghe si atentia.
SRIA nu este un sistem la fel de sigur ca SRAA
ns datele experimentale si clinice l-au convins pe
Leon Dnil (1933- ) de existenta acestuia, pe care l-a
Fig. 3.13. Dispunerea FR n interiorul trunchiului
cerebral (cf. Olszewski).
Curs de neuropsihologie 25
descris, n 1972. Lezarea SRIA duce la aparitia
sindromului de logoree si hiperkinezie. SRAA si SRIA
sunt reciproc conectate si actioneaz n mod continuu,
iar veghea si somnul, mpreun cu ntreaga gam de
aspecte intermediare curprins ntre ele, rezult din
competitia functional a celor dou sisteme (desigur, si
alte sisteme sunt implicate n aceste dou stri
sistemul endocrin, circulator, SNV, dar si functii
psihice motivatia, vointa, gndirea etc.).
SRD are rol n controlul tonusului muscular.
SRD primeste impulsuri de origine cortical, extrapira-
midal, cerebeloas si vestibular si faciliteaz (SRAD)
sau inhib (SRID) activitatea muscular, prin actiunea
sa asupra neuronilor din coarnele anterioare ale mdu-
vei spinrii (tractul reticulospinal). Prin urmare,
alertarea cortexului de ctre SRAA este urmat de
alertarea SRD de ctre cortex.
Neuronii FR sunt importante surse de secretii
ale unor neurotransmittori, cum ar fi : noradrenalina,
dopamina, serotonina si cel putin 40 de peptide.
n concluzie, FR este o substant nervoas care
controleaz starea de somn i de veghe, precum i toni-
fierea psihic i muscular.
3.2.5. Patologia trunchiului cerebral
Trunchiul cerebral este structura anatomic ce
contine cea mai mare parte a componentelor vitale din
ntregul SNC. Acesta contine nucleii nervilor cranieni si
fibrele acestora, tracturile senzoriale lungi ascendente si
descendente corticale si subcorticale, formatiunea
reticulat cu rol n activarea cerebral si apeductul cere-
bral (Sylvius), care conduce lichidul cerebrospinal.
Leziunile care afecteaz direct tesutul trunchiu-
lui cerebral se numesc intraaxiale sauparenchimale, iar
cele situate n afara trunchiului cerebral, ce afecteaz
meningele sau nervii cranieni, sunt numiteextraaxiale.
De asemenea, pot fi unilaterale si bilaterale. Efectele
unei leziuni pot fi ipsilaterale (pe aceeasi parte) sau
contralaterale (pe partea opus).
Un exemplu de leziune extraaxial este tumora
numit neuronimul de acustic. Acesta se dezvolt din
celulele Schwann ale nervului vestibulocohlear (VIII).
Mai nti, neurinomul afecteaz nervul acustic (VIII),
dup care ncepe s comprime (apese) trunchiul
cerebral, nervii cranieni din jur si cerebelul. Pacientul
semnaleaz pierderea auzului, diminuarea reflexului
cornean (de nchiderea a pleaopelor la stimulri puter-
nice sau nocive), parez facial periferic, ataxie (impo-
sibilitatea de a coordona miscrile precise) si tremor
intentional (la miscrile voluntare).
Si alte procese expansive mari de fos cranian
posterioar pot duce la aparitia simptomelor de trunchi
cerebral, fie prin comprimare, fie prin infiltrare secun-
dar a acestuia. Acestea pot fi : meningioame (tumor
benign care se dezvolt pe arahnoid), abcese cerebe-
loase, astrocitoame (tumor de diverse malignitti,
aprut din astrocite), ependinoamele (tumori dezvol-
tate din celulele ependimale) de ventricul IV etc.
Medicul trebuie s cunoasc specializarea
functional a fiecrei substructuri anatomice pentru a
corela simptomele cu zona afectat. Pentru aceasta
trebuie s cunoasc aferentele si eferentele care circul
prin bulb, punte si mezencefal, localizrile reale si
aparente ale nervilor s.a.m.d.
Majoritatea leziunilor bulbului si punii sunt
vasculare, motiv pentru care nu vom intra n detalii,
fiind necesare, dup prerea noastr, cunostinte despre
sistemul vascular cerebral pentru a ntelege aceste tipuri
de leziuni. Pentru pons, totusi, am prezentat neurinomul
de acustic.
Dintre sindroamele mezencefalului, amintim
urmtoarele :
- Sindromul Weber : paralizia nervului oculomotor
(III) prin lezarea fibrelor emergente ale nervului ;
- Sindromul tegmental central : paralizie de nerv
oculomotor, provocat de lezarea nucleului sau
fibrelor nervului, la care se adaug tremor sau mis-
cri involuntare contralaterale (hemicoree) atribui-
te lezrii nucleului rosu ;
- Sindromul Benedikt : paralizie ipsilateral de nerv
oculomotor si din miscri involuntare contralate-
rale ;
- Sindromul Claude : paralizie ipsilateral de nerv
oculomotor ;
- Sindromul Fowille peduncular : sindrom Weber
asociat cu paralizia privirii laterale, bolnavul
privind spre partea sntoas ;
- Sindromul nucleului rosu Foix : miscri involunta-
re si tulburri de sensibilitate, fr afectarea
nervului oculomotor ;
- Sindromul nucleului rosu de tip inferior (Benedikt)
paralizie ipsilateral de nerv oculomotor ;
- Sindromul Perinauld : paralizia miscrii de vertica-
litate a ochilor.
- Sindromul lui von Monakov (sindromul senzitiv al
calotei) : paralizia ipsilateral a nervului oculomo-
tor si aparitia hemiansteziei si hemiparesteziilor
contralateral ;
Curs de neuropsihologie 26
- Sindromul locusului niger (substanta neagr) :
tulburri de tonus muscular ;
- Sindromul loked-in : apare n cazul lezrii bilatera-
le a tectumului prin ocluzia arterei bazilare si duce
la ntreruperea bilateral a tracturilor corticobulbar
si corticospinal, care provoac tetraplegie (paralizi-
a tuturor membrelor), mutenie si parez facial.
Este o afectiune extrem de grav, putnd fi
confundat cu coma, ns bolnavul este constient,
are ochii deschisi si i poate misca voluntar, clipitul
si miscarea ochilor fiind singurele sale capacitti
de comunicare.
Este, desigur, foarte dificil a nvta toate aceste
sindroame de-o dat. De aceea, sintetizm si artm c
cele mai multe sindroame ce apar sunt legate de para-
lizia nervului oculomotor (Claude, Benedikt etc.), de
tonus muscular (sindromul locusului niger), diverse
forme de anestezii, paralizii si miscri involuntare
ipsilaterale si contralaterale (implicate, de regul, n
cadrul celorlalte dou mari categorii de sindroame).
n ceea ce priveste patologia formaiunii reti-
culate, vom vorbi despre dou sindroame : mutismul
akinetic si sindromul de logoree si hiperkinezie.
Mutismul akinetic (gr. kinetikos cauz a mis-
crii) este o tulburare permanent sau intermitent a st-
rii de constient, caracterizat prin lipsa de rspuns la
ntrebri (mutism) si imobilitate a capului si corpului
(akinezie). Bolnavul tine ochii deschisi, i deplaseaz
spre examinator, spre obiectele prezentate si spre sursa
de zgomot, dar nu are nici un rspuns emotional sau
motor la ncercrile examinatorului de a comunica cu el
si nici fat de tot ce se petrece n jur. Un aspect foarte
important este c bolnavul nu este paralizat, ci doar
incapabil de a reactiona. n cazul episoadelor intermi-
tente, bolnavul poate spune nume de persoane si fapte
prezentate sau petrecute n faza total de akinezie si
mutism. Exist, pe lng mutismul akinetic de origine
reticulat, si un mutism akinetic de origine frontal sau
cingular (cortexul frontal). Dup cum am artat mai
sus, reflexul de orientare este pstrat n ciuda imobilit-
tii totale, ritmul somn-veghe este pstrat si este posibil
alimentatia pasiv, deglutitia fiind n stare relativ bun,
dar, desigur, bolnavul nu poate exprima dac i place
sau nu ce mnnc. Se pare c mutismul akinetic este
provocat de lezarea SRAA.
Sindromul de logoree i hiperkinezie a fost
descris de Dnil si Arseni (1977), care au observat c
starea bolnavilor poate avea si un aspect opus mutismu-
lui akinetic, respectiv logoree (trncneal excesiv)
si hiperkinezie (hiperactivitate motorie) sau alte feno-
mene intermediare (logoree fr hiperkinezie, hiperki-
nezie fr logoree, hiperkinezie cu mutism). Acest sin-
drom apare att preoperator, ct si postoperator la bol-
navii cu diverse leziuni de tip tumoral sau vascular. Sin-
dromul se caracterizeaz prin hipermnezie (exagerarea
evocrii faptelor trecute), exagerarea atentiei, a
reflexului de orientare (bolnavul ntoarce capul si
tresare la o multime de stimuli nesemnificativi), mobili-
tate exagerat, simt critic ascutit, somn foarte putin (2-3
ore din 24). Bolnavul poate pstra rationamente
normale, n functie de ntinderea si localizarea leziunii.
Existenta acestui sindrom i-a condus pe cei doi cercet-
tori romni (Dnil, 1972, Studiu clinic i experimen-
tal asupra psihopatologiei substantei reticulate ;
Arseni, Dnil, 1977, Logorrhea syndrome with hyper-
kinesia) s afirme existenta SRIA, care este afectat de
acest sindrom.
Pe lng aceste sindroame, leziunea FR mai
poate conduce si la rigiditate a tonusului muscular,
hipertonicitate, dereglrii ale posturii corpului s.a.
3.2.6. O divizare alternativ a componente-
lor i histochimia trunchiului cerebral
1
n pofida unor diferente structurale ce apar ntre
medulla oblongata, pons si mezencefal, trunchiul cere-
bral are o structur uniform (fig. 3.14). Astfel, avem :
- Tegmentumul sau calota, partea cea mai veche
filogenetic, comun tuturor celor trei componente
si care contine nucleii nervilor cranieni. La nivelul
bulbului si puntii, este acoperit de cerebel, iar la
nivelul mezencefalului, este acoperit de tectum sau
lama cvadrigemen alctuit din coliculii cvadrige-
meni inferiori si superiori. Tegmentumul reprezint
partea posterioar a bulbului si puntii, dar la niveul
mezencefalului, acesta este partea de mijloc, cea
posterioar fiind reprezentat de tectum ;
- Tracturile descendente (motorii) constituie partea
anterior a trunchiului cerebral. Acestea formeaz
piramidele la nivelul bulbului, piciorul (bulbul
pontin, piramidele pontine) n pons si crus cerebri
(picioarele pedunculilor cerebrali) la nivelul
mezencefalului.
Regiunile trunchiului cerebral difer, de asemenea,
si n ceea ce priveste compozitia chimic. Delimitarea
1
Nepredate.
Fig, 3.14. Divizarea pe vertical a trunchiului cerebral.
Curs de neuropsihologie 27
ariilor n functie de structura chimic pe care o prezint
se numestechemoarhitectur.
S-a descoperit c trunchiul cerebral continefier,
n special n substanta neagr si, ntr-o msur mai
mic, si n nucleul rosu (prezenta fierului aici se face, n
parte, responsabil pentru culoarea aproape rosie a
acestui nucleu). Fierul se gseste n neuroni si n
celulele gliale, sub form de mici particule, iar prezenta
fierului este o caracteristic a tractului extrapiramidal.
Neurotransmittorii i enzimele necesare pen-
tru sinteza acestora au urmtoarea chemoarhitectur :
neuroni catecolaminergici (secret norepinefrin, epi-
nefrin si dopamin) si serotoninergici se afl n nucleii
specifi ai tegmentumului, iar n nucleii nervilor cranieni
se gsesc neuroni colinergici. Analiza histochimic
relev prezenta unui continut bogat de norepinefrin n
tegmentumul mezencefalic si o cantitate semnificativ
mai mic n tectum si n tegmentumul bulbar.
Dopamina se gseste din plin n substanta neagr si
foarte putin n restul trunchiului cerebral.
Trunchiul cerebral prezint si o chemoarhitectu-
r a enzimelor metabolice. Activitatea enzimelor oxi-
dante este, n general, mai ridicat n substanta cenusie
(n special n nucleii nervilor cranieni, portiunea
inferior a olivelor si n nucleii pontuberantei). Distri-
buirea enzimelor este caracteristic fiecrei arii cenusii
si se foloseste denumirea depattern enzimal.
3.2.7. Nervii cranieni
Exist 12 perechi de nervi cranieni, din care 10
intr n trunchiul cerebral sau ies din acesta (n afar de
I si II). Majoritatea acestor nervi mediaz functiile
senzoriale sau motorii ale capului si gtului. Nervul vag
(X) controleaz functiile organelor din cavitatea
toracic si abdominal. Se numeste vag pentru c nu
inerveaz o zon precis, ramificatiile sale fiind
mprstiate prin cavittile toracic si abdominal. Unii
nervi sunt asociati numai cu functiile senzoriale (optic,
II), altii doar cu functiile motorii (hipoglos, XII), iar
altii pot avea o combinatie de functii. Acestea pot fi :
senzoriale si motorii (trigemen, V), senzoriale, motorii
si vegetative (glosofaringean, XI) sau motorii si
vegetative (oculomotor, III).
Mai multe detalii despre nervii cranieni se
gsesc n tabelul 3.1., iar proiectiile fiecruia se pot
observa n fig. 3.15.
Fig. 3.15. Emergena nervilor cranieni, modalitile senzoriale pe care le conduc i efectorii pe care i inerveaz.
Curs de neuropsihologie 28
Tabelul 3.1. Nervii cranieni. Tipuri, funcii, emergena din trunchiul cerebral i patologie.
I Olfactiv
Mediatorul modalittii senzoriale olfactive (miros), singura cale senzorial care nu face
sinaps precortical, n talamus. Este un nerv visceral aferent alctuit din axonii nemielinizati
ai neuronilor bipolari din mucoasa nazal (epiteliul olfactiv).
II Optic
Nerv special somatic aferent care mediaz vederea si reflexele pupilare la lumin. Un nerv
optic sectionat nu se reface (orbire).
III Oculomotor
Nerv somatic eferent visceral care misc globul ocular, strnge pleoapele, acomodeaz globul
pentru vedere optim. Iese din trunchiul cerebral prin santul interpeduncular al
mezencefalului. Patologie: paralizia oculomotorie, atrnarea pleaopei (ptosis), pupil dilatat,
fix.
IV Trohlear
Nerv somatic general eferent care inerveaz muschiul oblic superior (miscri oculare). Iese,
contralateral (din stnga pentru ochiul drept si invers), din partea dorsal a mezencefalului.
Patologie: rigidate a miscrilor oculare, diplopie (veder dubl) vertical s.a.
V Trigemen
Nerv special visceral eferent (iese din punte si inerveaz muschii masticatiei) si somatic
aferent general (la nivelul fetei, mucoaselor cavittii bucale si nazale, sinusului frontal si
structurile profunde ale capului). Patologie: hemianestezie facial si a mucoasei cavittii
nazale si bucale, pierderea reflexului corneean, paralizia muschilor masticatori, devierea flcii
pe partea afectat, nevralgie trigeminal (durere acut junghi repetat n una sau mai multe
ramuri ale nervului).
VI Abducens
Nerv somatic eferent general care inerveaz muschiul drept lateral al ochiului. Porneste din
nucleul abducens aflat n partea posterioar a puntii. Patologie: hemipareza abducens,
strabism convergent, diplopie orizontal.
VII Facial
Nerv somatic aferent general (inerveaz suprafata posterioar extern a urechii), visceral
aferent general (palat si peretii faringelui), visceral aferent special (mugurii gustativi din
partea anterioar a limbii), visceral eferent general (glandele lacrimale, submandibulare,
sublinguale) si visceral eferent special (muschii expresiilor faciale). Patologie: paralizie
facial, pierderea reflexului corneean, pierderea functiei lacrimale, uscciunea gurii, pierderea
gustului (posterior).
VIII Vestibulocohlear
Nerv somatic aferent special cu dou subdiviziuni functionale: unnerv vestibular (mentinerea
echilibrului si balansului) si unul cohlear (mediaz auzul). Iese din partea dorsal a puntii.
Patologie: vestibularul dezechilibru, ameteal, miscarea involuntar a ochilor; cohlearul
surzenie, neurinomul de acustic.
IX Glosofaringean
Nerv somatic aferent general (inerveaz o parte a urechii externe), visceral aferent general
(mucoasa cavittii bucale s.a.), visceral aferent special (inverveaz mugurii gustativi din 1/3
posterioar a limbii), visceral eferent special (ridicarea faringelui n timpul deglutitiei),
visceral eferent general (inerveaz glanda parotid care elimin saliv). Patologie: disfagie,
pierderea gustului n partea posterioar, anestezie, pierderea reflexului faringean.
X Vag
Nerv somatic aferent general (urechea extern si membrana timpanului), visceral aferent
general (faringe, laringe, esofag, trahee, torace, abdomen), visceral aferent special (muguri
gustativi din regiunea epiglotic), visceral eferent special (muschii faringelui, laringelui,
esofagului etc.), visceral eferent general (interiorul gtului, toracelui si stomacului).
Patologie: disfonie (cauzat de paralizie ipsilateral a faringelui si laringelui), disfagie,
devierea boltei palatine, pierderea reflexului de tuse
XI Accesoriu
Nerv visceral eferent special (mediaz miscrile capului si umerilor si inerveaz muschii
larigeali), mprtit n diviziunea accesorie (cranian) iese din bulb si spinal iese din
coarnele ventrale al C1-C6. Patologie: paralizia muschiului sternocleidomastoidian (dificulate
n ntoarcerea capului pe partea opus), paralizia muschiului trapezoid (lsarea umerilor),
paralizia laringelui.
XII Hipoglos
Nerv somatic eferent general: mediaz miscarea limbii prin inervarea muschilor interni si
externi ai acesteia. Iese din bulb. Patologie: hemipareza limbii, protruzia limbii pe partea
afectat.
Tipuri de nervi
Afereni
General somatic aferent Nerv care mediaz sensibilitatea cutanat (de durere, atingere si temperatur).
Special somatic aferent Nervcare mediaz sensibilittile speciale (vz, auz, miros, gust).
Special visceral aferent Nerv care mediaz sensibilittile chimice (gust, miros).
General visceral aferent Nerv care mediaz sensibilittile interoceptive (foame, sete, durere organic).
Efereni
General somatic eferent Nerv care inerveaz muschii scheletici.
Special visceral eferent Nerv care inerveaz muschii derivati din arcul faringean (ai masticatiei).
General visceral eferent Inerveaz muschii netezi sau glandele.
Curs de neuropsihologie 29
3.3. CEREBELUL
n partea posterioar a trunchiului cerebral, putin deasu-
pra bulbului rahidian si acoperit de lobii occipitali, se
afl cerebelul (lat. cerebelus cereier mic, creieras), o
structur anatomic de form oval, cu aspect striat
(fig. 3.9). Cerebelul apare destul de trziu din punct de
vedere filogenetic, ntlnindu-se abia la reptile. Denu-
mirea de creieras se datoreaz aspectului su, destul de
asemntor cu cel al creierului mare (cerebrum).
Asemnrile dintre cerebel si cerebrum constau n: lipsa
emergentelor nervilor cranieni, divizarea suprafetei n
santuri si circumvolutii, tipul de structura intern, exis-
tenta fibrelor de proiectie, conexiuni corticosubcorticale
si altele. Diferentele se refer la ngustimea circumvolu-
tiilor, la dispunerea mai ordonat a acestora la cerebel si
la uniformitatea structurii corticale a cerebelului. Spre
deosebire de scoarta cerebral, unde celula predominan-
t este cea piramidal (neuronul motor), n scoarta cere-
beloas, dominant este celula Purkinje.
Cerebelul (ca si cerebrum) are dou emisfere.
Aceste emisfere sunt mprtite n trei lobi (fig. 3.16) :
lobul anterior (situat cel mai aproape de trunchiul
cerebral), lobul posterior (cel mai mare, delimitat de cel
anterior prin fisura primar) si lobul flocculonodular
(cel mai mic, asezat caudal si separat de lobul posterior
prin fisura posterolateral). Din punct de vedere
filogenetic si functional, cerebelul se divide n :
arhicerebel (lobul flocculonodular), paleocerebel
(spinocerebel) si neocerebel (cerebrocerebel).
Vermisul este o structur ce prezint circumvo-
lutii transverse inelate (de unde si ia si numele de
vermis, dat de Galen) situat ntre cele dou emisfere
cerebeloase (fig. 3.17). Pe suprafata vermisului si pe
suprafata emisferelor se pot distingelobuli (mici lobi).
Fiecare lobul de pe vermis se continu cu unul de pe
emisfere, alctuind o unitate morfofunctional. Un
exemplu de lobul esteamigdala
1
(fig. 3.17).
Structura intern a cerebelului prezint sub-
stant cenusie dispus la exterior pentru a forma scoarta
cerebeloas, dar si n profunzime, sub forma unor
nuclei (nucleii cerebeloi profunzi) nconjurati de
substant alb.
Scoara cerebeloas sau cortexul cerebelos
prezint o organizare neuronal stratificat (fig. 3.18).
Acestea sunt : stratul molecular, stratul ganglionar
(Purkinje) si stratul granular si contin cinci tipuri de
neuroni : stelati, n form de cos, Purkinje, Golgi si
celulele granulare.
1
A nu se confunda cu nucleul amigdaloid din sistemul limbic
(cap. 3.5.1.-2).
Fig. 3.16. Localizarea cerebelului i a lobilor cerebeloi.
Fig. 3.17. Cerebelul vzut de sus (A) i de jos (B).
Curs de neuropsihologie 30
1) Stratul molecular este situat la exterior si este
populat, n principal, cu celule cu form de cos si
celule stelate. Axonii si dendritele celulelor stelate
nu prsesc acest strat, precum nici dendritele celu-
lelor n form de cos. n acest strat se mai gsesc si
dendritele neuronilor Purkinje din stratul Purkinje,
axonii celulelor granulare si ai celulelor Golgi ;
2) Stratul Purkinje (ganglionar, intermediar) este
alctuit, desigur, din celule Purkinje, niste
celule tipice cerebelului. Acesti neuroni au di-
mensiuni mari (50-80 mm), au form de par
si sunt asezate unele lng altele ntr-un singur
strat. Axonii celulelor Purkinje reprezint sin-
gurele fibre eferente ale cortexului cerebelos.
Toti neuronii Purkinje sunt inhibitori, folosind
neurotransmittorul GABA (vezi cap. 2.4.).
3) Stratul granular (intern) contine celule granu-
late si celule Golgi. Dendritele celulelor granu-
late au form de ghear, iar axonii lor si axonii
neuronilor Golgi sunt orientati n sus, ajun-
gnd pn la stratul molecular. n acest strat,
se mai gsesc si glomeruli, mici spatii unde
neuronii stratului granular fac sinaps cu
fibrele muchiulare
1
.
1
n general poart denumirea de ramificate. Prof. Dnil
si Golu le numesc muschiulare deoarece au forma muschiului
de copac, iar noi vom folosi n continuare aceast denumire.
Exist dou tipuri deaferente ctre scoarta ce-
rebeloas. Primul este cel de care am vorbit, care se rea-
lizeaz la nivelul glomerulilor (prin intermediul fibrelor
muschiulare). Cel de-al doilea tip de aferente se reali-
zeaz prin intermediul fibrelor ctrtoare care fac si-
naps cu celulele Purkinje, acestea din urm transmi-
tnd impulsul mai departe, la nucleii cerebelosi.
Funciile scoarei cerebeloase. Sunt legate, n
principal, de functionarea neuronilor Purkinje, care
contin singurii axoni eferenti din scoart. Activitatea
neuronilor Purkinje este determinat de dou aferente
excitatorii : fibrele muschiulare si fibrele ctrtoare.
Fibrele ctrtoare au originea la nivelul nucleilor
olivari din bulbul rahidian, iar fibrele muschiulare
provin din regiunile cerebrale superioare, trunchiul
cerebral i mduva spinrii.
Singurul neuron excitator de la nivelul
cortexului cerebelos este neuronul granular. Neuronii
stelati si cei n form de cos inhib neuronii Purkinje,
iar neuronii Golgi inhib celulele granulare. Eferentele
neuronilor Purkinje inhib neuronii nucleilor cerebelosi,
acestia din urm dnd nastere fibrelor eferente ale
cerebelului. Prin urmare, principalul rol al scoartei cere-
beloase este de a regla fin, prin mecanismele de inhibi-
tie, eferentele nucleilor profunzi cerebelosi.
Se presupune, de asemenea, c acesti neuroni
sunt implicati si n unele forme de nvtare motorie.
Nucleii cerebeloi profunzi sunt situati n pro-
funzimea cerebelului, fiind nconjurati de substanta
Fig. 3.18. Structura intern a cerebelului.
Curs de neuropsihologie 31
alb. Acesti nuclei se gsesc, sub form de perechi, att
n vermis, ct si n emisferele cerebeloase, si primesc
aferente de la dou surse : 1) cortexul cerebelos (neu-
ronii Purkinje) si 2) tracturile aferente cerebeloase
(ctrtoare si muschiulare), care provin din mduva
spinrii, trunchiul cerebral si structurile cerebrale
superioare. Exist trei tipuri de nuclei cerebelosi
profunzi :
1) Nucleii fastigiali : situati n vermis si au legturi
aferente si eferente cu nucleii vestibulari din bulbul
rahidian si cu formatiunea reticulat ;
2) Nucleii globulari sau interpui : situati n emisfere-
le cerebeloase unul n stnga, altul n dreapta ,
lateral de nucleii fastigiali, trimit aferente ctre
nucleul rosu (din mezencefal) si talamus (nucleul
ventrolateral) ;
3) Nucleii dintati sau dantelati : situati lateral de nu-
cleii globulari, aproximativ n centrul emisferelor
cerebeloase, cei mai mari si trimit eferente ctre
nucleul rosu, talamus si globus pallidus (compo-
nent a ganglionilor bazali).
Substana alb a cerebelului este format din
fibre mielinizate, care din punct de vedere functional se
mpart n :
- Fibre de proiectie : sunt cele mai lungi si asigur
comunicare cerebelului cu celelalte componente
ale SNC ;
- Fibrele comisurale : sunt relativ scurte, ele asigu-
rnd comunicarea ntre zonele simetric opuse ale
emisferelor cerebeloase (intermisferic) ;
- Fibrele de asociere : sunt foarte scurte, deoarece
asigur comunicarea intraemisferic (ntre dou
locatii din aceeasi emisfer) sau de la scoarta cere-
beloas la nucleii cerebelosi profunzi.
Legturile cerebelului ce restul SNC se fac prin
intermediul celor trei perechi de fibre nervoase mari nu-
mitepedunculi cerebeloi (fig. 3.17). S-i analizm pe
rnd.
1) Pedunculii cerebeloi superiori (brachium conjun-
ctivum) fac legtura ntre cerebel si mezencefal,
contin mai multe fibre eferente dect aferente, fibre
care pornesc, n mare parte, din nucleii dintati si
ajung la mduva spinrii, trunchiul cerebral, tala-
mus si scoarta cerebral.
2) Pedunculii cerebeloi mijlocii (brachium pontis)
leag cerebelul de punte, sunt cei mai voluminosi,
contin fibre pontocerebeloase, comisurale, de
asociere si corticocerebeloase (leag scoarta cere-
bral de cerebel) ; ei mediaz impulsurile miscri-
lor voluntare de la cerebrum (creierul mare), care
trec prin punte si ajung la cerebel.
3) Pedunculii cerebeloi inferiori (corpi restiformi)
contin fibre aferente (vestibulare si proprioceptive)
si eferente (motorii) spre mduv si bulb.
Funciile cerebelului sunt legate, n principal,
de activitatea motorie, iar mai nou, se crede c ar avea
si un rol cognitiv, n nvtarea motorie. Cele trei functii
clasice ale cerebelului (Gordon Holmes, 1930) sunt :
coordonarea activittii motorii, controlul tonusului
muscular si mentinerea i controlul echilibrului.
Cerebelul contribuie la executarea micrilor
fine, precise i voluntare (care tin n special de apucare
si manipulare a obiectelor, cum ar fi desenatul, de ex.).
Impulsurile pentru aceste miscri pornesc din scoarta
cerebral si sunt coordonate de ctre cerebel. Din acest
punct de vedere, putem vedea functiile cerebelului ca pe
un ajutor dat scoartei cerebrale, menit s integreze
toate informatiile legate de miscare si postur.
Fiecare miscare are la baz actiunea coordonat
a unor grupe musculare. Cerebelul are rolul de a genera
rapid, la debutul unui act motor, impulsuri nervoase de
tip pornit pentru muschii agonisti (care provoac mis-
carea) si, simultan, impulsuri nervoase de tip oprit
pentru muschii antagonisi (care se opun efectului mus-
chilor agonisti). Pentru finalizarea miscrii, cerebelul va
transmite impulsuri de oprit pentru agonisti si
impulsuri de pornit pentru antagonisti. Se poate dedu-
ce de aici c o coordonare rapid si alternativ a acestor
impulsuri faciliteaz miscrile fine voluntare.
Cerebelul este un centru integrativ important.
Aceasta nseamn c el primeste aferente de la toate
regiunile SNC asociate cu miscarea (mduv, trunchi,
talamus, scoarta cerebral), prelucreaz semnalele si tri-
mite napoi unfeedback. De asemenea, se presupune c
ar contine un mecanism de modelare a transmiterii
impulsurilor nervoase senzoriale ctre scoarta cerebra-
l (proprioceptive, vestibulare si tactile) si c ar avea un
rol dereglare a functiilor vegetative ale organismului.
Unii cercettori avanseaz ipoteza c cerebelul ar avea
un rol si ninitierea cuvntului i a limbajului.
Cercetrile experimentale arat c cerebelul
este implicat nnvtarea motorie. Responsabili sunt, n
principal, neuronii Purkinje.
Exist trei arii functionale cerebeloase cu rol n
controlul motor. Astfel, avem :
- Vestibulocerebelul (lobul flocculonodular) contro-
leaz miscrile corpului cu rol n mentinerea echi-
librului ;
- Spinocerebelul (paleocerebelul) coordoneaz mis-
crile segmentelor distale (deprtate) ale membre-
lor, labele, minile si degetele ;
Curs de neuropsihologie 32
- Cerebrocerebelul (neocerebelul) are legturi de tip
feedback cu scoarta cerebral pentru planificarea
miscrilor voluntare ale corpului si membrelor
(proces denumit formarea imaginii motorii).
Patologia cerebelului. Pentru a produce o
disfunctie grav i permanent a cerebelului, leziunea
trebuie s implice cel putin unul din cei trei nuclei cere-
beloi profunzi (dintat, globular sau fastigial). O leziune
cerebeloas produce afectri ipsilaterale ale membrelor.
De cele mai multe ori, doar o pereche de membre este
afectat, iar afectare membrelor inferioare este mai
putin rspndit. Pacientii sufer, n general, si de orto-
statism (dificultate n a sta n picioare).
Cerebelul se vindec, sau functiile sale sunt
preluate de ctre scoarta cerebral. O leziune cerebe-
loas reaprut poate duce la disfunctii, dar acest caz
este mai rar, n general functiile fiind preluate, dup
cum am mai zis, de ctre scoarta cerebral.
Primele afectiuni cerebeloase au fost descrise
de ctre Gordon Holmes (1876 1966), ntre anii 1920
si 1930. Autorul a descris prezenta a trei deficite
principale la pacientii cu plgi cerebeloase provocate de
glont, n primul rzboi mondial. Dup cum am precizat
anterior, acelasi neurolog a determinat si functiile
clasice ale cerebelului, n functie de aceste trei
afectiuni. Acestea sunt :
- Hipotonia este datorat pierderii functiei de facili-
tare pe care cerebelul o exercit asupra scoartei
cerebrale si asupra trunchiului cerebral, care deter-
min scderea tonusului musculaturii membrelor,
ipsilateral leziunii. Sunt afectati n special nucleul
dintat si cel globular. Bolnavul manifest pierderea
rezistentei la miscri pasive, ntrzierea rspunsu-
lui de efectuare a unor miscri rapide, inabilitatea
opririi rapide a miscrii unui membru si reflexe
pendulare (atrnarea membrelor).
- Ataxia se reflect, n principal, prin erori n efec-
tuarea miscrilor (dismetrie), de exemplu, dac
bolnavului i se cere s-si ating nasul cu mna, nu
va reusi, atingnd o portiune de deasupra sau de
dedesuptul nasului. Lezarea cerebelului face ca
scoarta cerebral s nu poat anticipa ct de ample
trebuie s fie miscrile. n consecint, miscrile de-
psesc, de obicei, limita intentional, iar cortexul
cerebral transmite impulsuri nervoase n exces
pentru efectuarea, n mod constient, a unei miscri
compensatorii.
- Tremorul intentional miscrile voluntare ale unei
persoane cu leziune cerebeloas sunt inexacte,
oscilante, n special atunci cnd se apropie de tinta
intentional. Se mai numestetremor de actiune si
este cauzat de de incapacitatea cerebelului de a
atenua miscrile, de a le da precizie.
Pe lng aceste afectri, au mai fost descoperi-
te si alte sindroame, cum ar fi : disartria (vorbire
dezorganizat, n care unele silabe sunt pronuntate
puternic, altele slab, altele prelungite si altele scurte,
astfel nct vorbirea devinte neinteligibil), nistagmus
cerebelos (tremorul globilor oculari la ncercarea de a
fixa privirea undeva n lateral) si disdiadocochinezia
(progresia ordonat a miscrilor nu se poate realiza).
3.4. DIENCEFALUL
Diencefalul este o structur anatomic major, situat
deasupra si n continuarea mezencefalului, dedesuptul
telencefalului si n jurul ventriculului III (fig. 3.9, fig.
3.5). Limita posterioar este dat de comisura poste-
roar si de corpii mamilari, limita anterioar este dat
de chiasma optic, iar limitele laterale sunt date de
bandetele optice, coada nucleului caudat, stria terminal
si bratul posterior al capsulei interne (fig. 3.19 ; se va
vorbi despre aceste componente n unittiile urmtoare).
n cele mai multe tratate, diencefalul este mprtit n
dou mari diviziuni : talamus si hipotalamus (fig. 3.9).
Pe lng acestea, diencefalul mai contine si
metatalamus, epitalamus si subtalamus
1
. n aceast
formul, diencefalul devine compus din talamencefal
(meta-, epi-, subtalamus si talamus) si hipotalamus.
1
Meta deasupra, epi n spate, sub dedesupt.
Curs de neuropsihologie 33
3.4.1. Talamusul
Talamusul (gr. thalamos camer luntric) re-
prezint partea dorsal a diencefalului (motiv pentru
care se mai ntlneste n literatura de specialitate cu
numele de talamus dorsal). Este situat aproape n mijlo-
cul emisferelor cerebrale, fiind, prin urmare, acoperit n
totalitate de acestea, iar n partea inferioar este acoperit
de mezencefal. n raport cu ventriculul III, talamusul se
afl pe fetele laterale ale acestuia (fig. 3.19). Este
desprtit de hipotalamus prin antul hipotalamic, ce
trece prin ventriculul III.
Talamusul este alctuit din doi corpi ovoizi
(identici, cte unul de fiecare parte, fig. 3.19, 3.20) de
materie cenusie, care se mai numesc si corpi optici, cu
toate c ei reprezint unmecanism de releu pentru toate
sensibilittile n afar de cea olfactiv. Corpii talamici
sunt legati prin adeziunea intertalamic (fig. 3.20).
Talamusul este conectat masiv si reciproc cu cortexul
cerebral (adic trimite eferente si primeste aferente de
la cortex). De asemenea, dispune si de conexiuni cu
nucleii subcorticali. Extremitatea posterioar este mai
voluminoas dect cea anterioar (fig. 3.20), este
rotunjit si ptrunde n ventriculii laterali (fig. 3.19). Se
mai numeste si pulvinar.
Substana cenuie a talamusului este mprtit
n cinci grupe denuclei (fig. 3.20), ce si iau numele n
functie de localizarea spatial, dup cum urmeaz (dup
Walker, 1930) :
1. Grupul nuclear anterior (nucleii anteriori si lateral
dorsal) este separat de restul talamusului prin lama
medular intern (n afar de nucleul lateral dorsal,
care este n afara lamei). Primeste aferente de la
corpii mamilari (tractul mamilotalamic) si trimite
eferente ctre cortexul cingulat si cortexul cerebral;
2. Grupul nuclear median, localizat sub cptuseala
ventriculului III, se conecteaz cu hipotalamusul si
substanta cenusie periapeductal (mezencefal) ;
3. Grupul nuclear medial este situat intern de lama
medular intern, include majoritatea substantei
cenusii mediale si nucleul dorsal medial care trimi-
te eferente ctre cortexul frontal si cel motor ;
4. Grupul nuclear lateral, cuprins ntre lama medu-
lar extern si cea intern, se ntinde de la partea
anterioar pn la pulvinar (partea posterioar),
primeste aferente senzoriale de la piele si trimite
eferente ctre lobul parietal ;
5. Grupul nuclear posterior include pulvinarul si
corpii geniculati (laterali si mediali) ; pulvinarul
este o mas nuclear mare, conectat cu lobul tem-
poral si cel parietal.
Dup cum se poate observa, n fig. 3.20, grupe-
le nucleare sunt mprtite n anterior, medial si lateral,
aceasta fiind o grupare ntlnit mai ales n literatura
anglo-saxon. Nu este la fel de complex ca gruparea
noastr initial, ns este mai usor de retinut, deoarece
se bazeaz pe mprtirea talamusul n trei prti (lateral,
Fig. 3.19. Seciune transversal prin baza creierului ce
relev poziionarea talamusului i a altor componente
aflate n vecintate. Partea anterioar a creierului este cea
unde se afl coada nucleului caudat (detaliu topografic).
Fig. 3.20. Nucleii talamici. VA ventral anterior, LD
lateral dorsal, VL ventral lateral, LP lateral
posterior, VP ventral posterior, VPL ventral
posterolateral, VPM ventral posteromedial,
VI ventral intermedial.
Curs de neuropsihologie 34
medial, anterior) de ctre lamina medular intern
1
.
Exist si alte tipuri de grupri ale nucleilor talamici, pe
care, ns, nu le vom mentiona aici.
n functie de conexiunile cu scoarta cerebral,
se pot diferentia dou tipuri de nuclei talamici : nuclei
specifici si nuclei nesepecifici.
Nucleii specifici sunt n legtur cu diferite mo-
dalitti senzoriale si se proiecteaz pe arii corticale
specifice. Se mai numesc nuclei de transmitere si
fiecare primeste impulsuri de la nu mai mult de o
modalitate senzorial, pe care le proiecteaz ntr-o zon
cortical specific de la care primeste un impuls
recurent. De exemplu, nucleii din corpii geniculati
laterali primesc impulsuri vizuale pe care le trimit ctre
cortexul vizual primar, iar corpii geniculati mediali pri-
mesc impulsuri auditive, pe care le trimit ctre cortexul
auditiv primar. Informatia senzorial a nucleilor speci-
fici reprezint prima etap a perceptiei, urmnd ca
scoarta cerebral s dea detalii de finete.
Nucleii nespecifici trimit proiectii difuze ctre
scoarta cerebral, nu localizate precis, ca n cazul nucle-
ilor specifici. Sunt, n principal, nucleii intralaminari si
reticulari. Nucleii intralaminari au legturi reciproce si
organizate cu cortexul, au eferente ctre corpul striat si
intervin n fenomenele de trezire. Nucleii reticulari
tapeaz suprafata extern a talamusului. n general,
natura si functiile relatiilor nespecifice dintre talamus si
cortex sunt necunoscute. n orice caz, datele experimen-
tale si clinice arat c talamusul joac un rol capital n
functionarea creierului.
Trebuie mentionat c nu toate aferentele primite
de ctre talamus ajung la scoarta cerebral, unele din
acestea terminndu-se n nucleii talamici.
Pe lng eferentele corticale, talamusul dispune
si deeferente subcorticale (nucleii intramediali si ven-
triculari) si eferente asociative (nucleii laterali). Eferen-
tele subcorticale pleac spre sistemul limbic, hipotala-
mus si mezencefal si primesc aferente de la cerebel si
unele tracturi ascendente ale trunchiului cerebral.
Eferentele asociative fac legtura ntre nucleii talamici
si au putine aferente.
Portiunile majore desubstan alb ale talamu-
sului sunt reprezentate de radiatiile talamice, lama
medular intern si lama medular extern. Radiatiile
talamice (corona radiata) sunt fibre nervoase care
pornesc din talamus si ajung n cortexul cerebral,
trecnd prin capsula intern. Lamina medular intern
se bifurc n portiunea sa anterioar, mprtind
1
La curs a fost predat prima grupare, prin urmare, aceasta
trebuie nvtat.
talamusul (dup cum aminteam mai sus) n trei prti
(anterioar, medial, posterioar). n profunzimea ei se
gsesc nucleii intralaminari (nu este vizibil n nici o
figur), nuclei nespecifici. Lamina medular extern
este situat pe suprafata lateral a talamusului si contine
nucleul reticular, de asemenea nespecific.
Recapitulnd, funciile talamusului sunt, n
principal, cea de releu a cilor sensibilittii (n afar de
cea olfactiv) si cea de activare difuz a scoartei. Func-
tia de releu se realizeaz prin intermediul nucleilor
specifici (cei prezentati n fig. 3.20), iar functia de
transmitere difuz, nespecific a semnalelor nervoase se
realizeaz prin intermediul nucleilor nespecifici (intra-
laminar, reticular).
De asemenea, talamusul este o component a
ciruitului Papez al emotiei (vezi pag. 49). n acest
circuit, stimulii emotionali ajung la talamus si
hipotalamus, care mediaz modificrile endocrine si
viscerale si care primesc, n acelasi timp, impulsuri
corticale prin hipocamp. Prin urmare, talamusul are si o
functie legat de afectivitate. Prin raporturile sale cu
hipotalamusul, talamusul joac un rol i n mecanisme-
le motivatiei. n ce priveste perceptia, dup cum am mai
amintit, talamusul reprezint prima etap a integrrii
senzoriale care formeaz perceptia, la unele specii de
animale fiind si ultima. Datele clinice (vezi mai jos)
arat c exist influente ale talamusului si n ceea ce
privestememoria si limbajul.
Patologia talamusului. Principala afectare a
talamusului se numeste sindromul talamic (descris de
ctre Djrine si Roussy n 1906). Acesta este constituit
din mai multe simptome :
- Hemianstezie contralateral (lipsa senzatiilor pe o
jumtate din suprafata corpului, jumtatea opus
leziunii talamice) apare imediat dup o hemoragie
talamic. Sensibilitatea superficial (de la suprafa-
t) poate s nu fie afectat deoarece are o reprezen-
tare bilateral n talamus ;
- Hemiparez contralateral ;
- Durere central (a capului) foarte intens, de tip
arsur, care se poate simti ntr-un ntreg corp tala-
mic afectat. Este dezagreabil, persistent, apare n
perioada de ameliorare a hemoragiei sau infarctului
talamic si se accentueaz la stress si oboseal ;
- Tremor intentional datorat lezrii nucleului lateral
ventral sau a aferentelor sale (de la cerebel, de ex) ;
- Ataxia membrelor inferioare (imposibilitatea de a
efectua miscri precise cu picioarele) este legat de
tulburarea sensibilittii profunde ;
Curs de neuropsihologie 35
- Tulburri de gust, uneori.
Afazia talamic este un sindrom caracterizat
prin tulburri ale fluentei verbale (ncetinire) si tulburri
ale expresiilor faciale, voce slab, rspunsuri scurte,
debit ncetinit si sacadat, discurs uneori incoerent. Pre-
supune o afectare persistent a memoriei logice si a
capacittii de asociere. Spre deosebire de alte afazii, cea
talamic are o evolutie bun, n afar de ceea ce prives-
te memoria cuvintelor.
Sindromul de neglijent talamic implic att o
neglijent motorie, ct si una spatial. Neglijenta moto-
rie se manifest prin imposibilitatea bolnavului de a-si
utiliza o jumtate din corp perfect functional. Exist o
terapie prin stimulare verbal, care face s dispar doar
partial neglijenta.
Neglijenta spatial este legat n special de
analizatorul vizual. Se manifest printr-o deviere a pri-
virii spre dreapta si refuzul de a explora spatiul din
stnga. Bolnavul manifest si o perceptie eronat a cor-
pului, deficiente n orientarea spatial, probleme de tip
apraxie constructiv (dificultti n a pune lucrurile
mpreun), spontaneitate excesiv a limbajului (spune
lucruri nesemnificative din senin), un deficit de nvtare
ce priveste materialul verbal si cel vizual.
Dementa talamic
1
(Schuster, 1936 ; Stern,
1939) se manifest prin pierderea spontaneittii motorii
si a initiativei verbale, mormieli neinteligibile, apatie,
indiferent fat de propria persoan si fat de anturaj,
gesturi stereotipice, absenta oricrei reactii afective,
anumite forme de amnezie si dezorientare spatio-
temporal. Dementa talamic apare dup lezarea bilate-
ral a conexiunilor talamusului cu cortexul cerebral si a
circuitelor hipocampo-talamo-cingulare. Cauza cea mai
frecvent a dementei talamice este infarctul bilateral.
Tulburrile afective se pot observa n cadrul
manifestrilor motorii sau expresive. Dup cum am
artat, talamusul intervine n circuitul Papez al emotii-
lor, prin conexiunile sale cu sistemul limbic. Functia
talamusului n afectivitate pare a fi amplificarea sau
diminuarea intensittii emotiilor. Pentru ameliorarea
simptomelor se pot efectua interventii chirurgicale.
3.4.2. Metatalamusul
Metatalamusul este alctuit din corpii genicu-
lati (mediali si laterali) si ocup o pozitie subtalamic
posterioar (fig. 3.20). Majoritatea tratatelor nu vor-
1
Sindrom nepredat.
besc despre metatalamus, ci includ aceste dou perechi
de nuclei n structura talamusului (dorsal).
Corpii geniculati laterali se leag de coliculii
cvadrigemeni superiori (cap. 3.2.3.). La acest nivel se
afl centrul vizual primar, care, la animale, realizeaz
unele integrri finale, permitnd astfel diferentiere ntre
lumin si ntuneric si orientri n raport cu localizarea si
intensitatea stimulilor luminosi. La om, aici se termin
tracturile vizuale (nervul optic), iar eferentele formeaz
radiatii optice, care ajung la scoarta cerebral (mecanis-
mul de releu pentru sensibilitatea vizual). Pe lng
aceste radiatii, de la corpul geniculat lateral mai pleac
fibre cu rol reflex spre coliculii cvadrigemeni inferiori.
Corpul geniculat lateral se compune din dou prti :
1) partea ventral (cea mai veche filogenetic) si 2) par-
tea dorsal (nou, filogentic). Ambele prti primesc
aferente vizuale, dar numai partea dorsal trimite efe-
rente ctre scoarta cerebral.
Corpii geniculati mediali se leag de coliculii
cvadrigemeni inferiori, formnd, la acest nivel, sinaps
cile auditive (nervul vestibulocohlear) conduse prin
lemniscul medial (cap. 3.2.1.\2.\3.). Mai departe, radia-
tiile acustice ajung la scoarta cerebral (ariile auditive
din lobul temporal). Primesc si alte aferente de la
trunchiul cerebral.
3.4.3. Epitalamusul
Epitalamusul (talamusul din spate) se afl n
partea superioar a diencefalului, ntre spleniul corpului
calos si coliculii cvadrigemeni superiori. Este alctuit
dinglanda pineal sau epifiza (fig. 3.9, 3.11), comisura
posterioar (epitalamic) si aparatul habenular.
Glanda pineal (epiphysis cerebri ; numele de
pineal provine de la asemnarea cu un con de brad)
este asezat ntre coliculii cvadrigemeni superiori (fig.
3.11). Descartes credea c aici se afl spiritul animale-
lor, dar n ziua de azi se stie c are alte functii. La verte-
bratele inferioare este un organ fotosenzitiv (modific
ritmurile noapte-zi ale organismului, n functie de
lumina pe care o primeste) si are o influent, n functie
de anotimp, asupra gonadelor (hormonii sexuali). La
vertebratele inferioare, lumina ptrunde direct ctre epi-
fiz, iar la vertebratele superioare, aceasta ptrunde prin
ochi. Epifiza are o actiune antigonadotropic (face ca
maturizarea sexual s nu aib loc pn la pubertate),
fiind observat hipergonadism la copiii care o au lezat.
Este, prin urmare, important n copilrie, dup care se
calcific. Are, de asemenea, un rol important n cres-
Curs de neuropsihologie 36
tere, iar datorit conexiunilor cu hipofiza poate influen-
ta ntregul sistem endocrin.
Celulele epifizei se numesc pinealocite si sunt
celule care au functia de a transforma informatia lumi-
noas n informatie endocrin (ndeplinind astfel func-
tiile pomenite mai sus). Hormonul principal secretat de
ctre epifiz estemelatonina, al crei precursor este 5-
HT (serotonina). Concentratia de 5-HT este supus va-
riatiilor circadiene (zi-noapte) : ziua maxim, noaptea
minim. Secretia de melatonin scade continuu de-a
lungul vietii. Nu s-a demonstrat un efect antigonadotrop
al melatoninei, dar se presupune c exist. Alte roluri
ale melatoninei sunt :
- modularea excitabilittii cerebrale ;
- influentarea pozitiv a somnului ;
- efect inhibitor al tumorilor canceroase.
Aparatul habenular (habenula) contine sub-
stant cenusie, alctuit din nucleii habenulari (intern si
extern) si mult substant alb prin care se conecteaz
cu alte regiuni subcorticale (aferente de la lamina
medular intern a talamusului, substanta perforat
anterioar, nucleul amigdaloid si stria terminalis si tri-
mite eferente ctre coliculii superiori, nucleul rosu si
nucleul intepeducular din mezencefal). Datorit legtu-
rilor sale, habenula este un sistem-releu pentru impulsu-
rile olfactive, care sunt transmise ctre nucleii salivatori
din trunchiul cerebral. n acest fel, se presupune c
mirosul influenteaz aportul de hran dorit.
3.4.4. Subtalamusul
1
Subtalamusul este continuarea tegmentumului
mezencefalic si contine nuclei ai sistemului extrapira-
midal (zona incerta, nucleul subtalamic, globus
pallidus), motiv pentru care poate fi considerat zona
motorie a diencefalului. Limita superioar este dat de
talamus (dorsal).
Zona incerta are legturi cu talamusul, nucleul
subtalamic, hipotalamusul si cu cile piramidale. Se
presupune c este o statie de releu pentru fibrele des-
cendente ale globului palid.
Globus pallidus este situat lateral si este mpr-
tit de o lamel de fibre mielinizate ntr-un segment
lateral (extern) si unul medial (intern). Segmentele sunt
interconectate prin fibre si sunt, de asemenea, conectate
cu putamenul si nucleul caudat. Are legturi reciproce
1
Nepredat la curs.
cu nucleul subtalamic si primeste aferente (n segmentul
medial) din substanta neagr.
S-a demonstrat, n pofida cunostintelor prece-
dente, c lezarea pallidumului nu provoac boala
Parkinson. Eliminarea unilateral a pallidumului, la
pacientii cu boala Parkinson, elimin ntepenirea
muscular contralateral si reduce tremorul. Eliminarea
bilateral cauzeaz dereglri psihice (sindromul post-
comotional : iritabilitate, oboseal, dificultate n con-
centrare).
Nucleul subtalamic (corpul lui Luys) este un
nucleu lenticular biconvex, de culoarea cafelei cu lapte.
Primeste aferente de la globus pallidus (partea lateral)
pe calea fasciculului subtalamic si trimite eferente tot
ctre globus pallidus (partea caudal a segmentului
medial). Unii autori au constatat legturi reciproce cu
talamusul, nucleul rosu, substanta neagr si
hipotalamusul.
La om, leziunile nucleului subtalamic dau
nastere unor miscri involuntare violente si persistente
ale bratului contralateral sau chiar hemibalism (misca-
rea jumttii corpului). Acest sindrom poart numele de
sindromul corpului lui Luys. La maimute, se obtine ace-
lasi sindrom prin distrugerea nucleului subtalamic.
Funciile subtalamusului (dup cum s-a putut
deduce din simptomele leziunilor nucleare) sunt n
legtur cu motricitatea. Nucleul subtalamic, mpreun
cu conexiunile sale cu corpul striat si tegmentumul
mezencefalic, reprezint locul de integrare al centrilor
de control motor.
3.4.5. Hipotalamusul
Cea de-a doua component major a diencefa-
lului, hipotalamusul, este asezat la baza creierului,
dedesuptul talamusului, nspre partea anterioar a
acestuia (fig. 3.9, fig. 3.21). Hipotalamusul este o
structur, am putea spune, fascinant (dar ce parte a
SNC nu este?), dat fiind faptul c ocup mai putin de
1% din suprafata creierului si controleaz nenumrate
functii ale organismului. Este singura portiune a
diencefalului care apare la suprafat, n zona mijlocie a
creierului. Este desprtit de talamus prin antul
hipotalamic, care constituie limita superioar si
posterioar a hipotalamusului. Limita anterioar este
dat de lamina terminal (fig. 3.22) - marginea
anterioar a ventriculului III. Poate fi mprtit n :
hipotalamus puternic mielinizat (corpii mamilari) si
hipotalamus slab mielinizat (restul hipotalamusului).
Curs de neuropsihologie 37
De asemenea, mai poate fi mprtit n : portiu-
nea supraoptic (anterior, deasupra chiasmei optice,
locul n care se ncruciseaz partial fibrele nervului
optic), portiunea tuberal (dorsal de portiunea
supraoptic) si portiunea mamilar (posterior, deasupra
corpilor mamilari). ntre chiasma optic si comisura
anterioar se gsestearia supraoptic (fig. 3.21).
Rostral de corpii mamilari (form de sn), apare
o zon bombat n directie inferioar, numit tuber
cinereum. n partea cea mai de jos a tuber cinereum se
prindetija pituitarei sau infundibulum, de care se leag
glanda pituitar (hipofiza) (fig. 3.21, 3.22).
Hipotalamusul este compus din 22 de nuclei
mici (substant cenusie), multe fibre nervoase care-l
traverseaz n lung si n lat (substant alb) si glanda
pituitar.
Nucleii hipotalamici sunt dispusi sub form de
mozaic si sunt destul de greu de identificat (fig. 3.21).
Fiecare nucleu hipotalamic ndeplineste o anumit
functie. Se mpart, de obicei, n patru grupe :
1. Grupul nuclear anterior (nucleul paraventricular si
nucleul supraoptic) secret hormoni care sunt
transportati la hipofiz (fig. 3.22) ;
2. Grupul nuclear lateral (nucleii tuberali laterali si
nucleul tuberomamilar) ;
3. Grupul nuclear mijlociu (nucleul ventromedial,
nucleul dorsomedial si nucleul infundibular sau
arcuat) ;
4. Grupul nuclear posterior (nucleii mamilari :
medial, care formeaz proeminenta corpului
mamilar fig. 3.21 si lateral).
Conexiunile hipotalamusului cu celelalte com-
ponente ale SNC, ct si cu sistemul endocrin prin
intermediul hipofizei, fac din acesta o structur
nervoas de important crucial pentru functionarea
sistemului psihic si a organismului. Hipotalamusul
dispune de conexiuni interne (intermediare) si
conexiuni externe (cu celelate componente ale SNC).
Este o component intermediar ntre talamus si scoarta
cerebral si ntre talamus si SNV.
Principalele ci aferente ale hipotalamusului
provin :
- de la sistemul limbic (mai specific, de la nucleul
amigdaloid prinstria terminalis si de la hipocamp
prinfornix) (fig. 3.21, fig. 3.22) ;
- pe calea fasciculului telencefalic medial (infor-
matii de la scoart si alte structuri subcorticale ;
scoarta are efect excitator si inhibitor asupra
hipotalamusului ; pe aceast cale mai ajung si
semnale olfactive si din principalii centri visce-
romotori ai trunchiului cerebral) ;
- de latalamus (nucleii mediali si anteriori) si de la
subtalamus (tractul palidohipotalamic).
- de la trunchiul cerebral (tegmentul mezencefalic,
lemniscul medial, substanta reticulat si substanta
alb).
Fig. 3.21. Seciune sagital prin crier ce relev localizarea principalilor nuclei talamici, a unor tracturi, a hipofizei, precum i
a altor componente ale SNC aflate n apropiere de hipotalamus sau cu care are legturi acesta.
Curs de neuropsihologie 38
- de la globii oculari (fibre retionotalamice) prin
rdcina chiasmei optice ; datorit acestor legturi,
semnalele luminoase pot influenta sistemul vege-
tativ (ritmul circadian si somn-veghe) si endocrin.
Cele mai importante ci eferente ale
hipotalamusului sunt :
- tractul hipotalamicohipofizar (fasciculul supraop-
ticohipofizar, paraventriculohipofizar si tubero-
infundibular) ;
- tractul hipotalamicohabenular (legturi cu
epitalamusul si, implicit, cu epifiza) ;
- tractul mamilotegmental se ndreapt ctre subs-
stanta cenusie din mezencefal (cea periapeductal)
si ctre nucleul interpeduncular ;
- tractul mamilotalamic (fig. 3.21) care se
proiecteaz pe grupul nuclear anterior ;
- fasciculul longitudinal dorsal (fig. 3.21) care
trimite fibre ctre coliculii cvadrigemeni, la toti
nucleii vegetativi ai trunchiului cerebral si la
formatiunea reticulat ;
- tractul tuberohipofizar ce contine fibre care pleac
de la portiunea tuberal si ajung la neurohipofiz.
- calea amigdaloid ventral care contine fibre
eferente (dar si eferente) ctre nucleul amigdaloid.
Glanda pituitar sau hipofiza (fig. 3.21, 3.22)
este un organ de dimensiuni mici (diametru de aproxi-
mativ 1 cm) si o greutate de 0,5-1 grame situat ntr-o
cavitate osoas de la nivelul bazei craniului numit
aua turceasc. Functiile hipofizei sunt strns legate de
conexiunile cu hipotalamusul, ce se realizeaz prin
intermediul tijei pituitare sau infundibulum. Din punct
de vedere fiziologic, hipofiza poate fi mprtit n dou
regiuni distincte : hipofiza anterioar (adenohipofiza
sau lobul anterior al glandei pituitare) si hipofiza
posterioar (neurohipofiza sau lobul neural al hipofi-
zei), dup cum se poate vedea n fig. 3.22. ntre cele
dou zone ale hipofizei se afl o arie de dimensiuni re-
duse (aproape inexistent la om), numit pars
intermedia (regiune intermediar).
Fiziologia hipotalamusului. Desi hipotalamu-
sul este o structur relativ mic, este una destul de
important. Putem gsi, ca o butad (poate cam piperat
pentru un curs de neuropsihologie), n literatura anglo-
saxon, afirmatia c hipotalamusul controleaz cei
patru F : fighting, feeding, fleeing and mating (lupta,
hrnirea, fuga si mperecherea). Acesti patru F reprezin-
t comportamentele necesare pentru supravietuirea
speciei.
n linii mari, hipotalamusul realizeaz controlul
homeostaziei. Termenul de homeostazie este folosit de
ctre fiziologi pentru a desemna mentinerea aproxima-
tiv constant a parametrilor mediului intern. Practic,
ntregul creier (de fapt, toate organele si tesuturile
organismului, mai mult sau mai putin) este implicat n
homeostazie, dar neuronii care o controleaz direct sunt
concentrati n hipotalamus. Prin intermediul hipotala-
musului, mediul intern al corpului este reglat de ctre
trei mari sisteme : 1) sistemul endocrin (SE) ;
2) sistemul nervos vegetativ ; 3) sistemul motivational.
1. Controlul SE este realizat de ctre
hipotalamus prin intermediul hipofizei. Eliberarea
majorittii hormonilor hipofizari se afl sub controlul
hipotalamusului, pe care acesta l exercit fie prin me-
canisme hormonale, fie prin semnale nervoase, motiv
pentru care hipofiza si reduce la un nivel extrem de
mic secretiile dac este mutat din locul ei de sub
hipotalamus. Controlul endocrin este mediat de ctre
dou clase de neuroni peptidergici neuroendocrini :
parvocelulari (cu celul mic) si magnocelulari (cu
celul mare). Fiecare neuron secret mai mult de o
peptid.
Gradul de activitate al acestor neuroni
particulari ai hipotalamusului depinde de stimulii
neurali dinuntrul si din afara organismului, precum si
de activitatea hormonal a variatelor organe tint
(tiroida, suprarenala, ovarele).
Neuronii neuroendocrini parvocelulari (din
nucleul preoptic, paraventricular, arcuat, tuberal, regiu-
nea bazal medial) sintetizeaz si secret hormoni
eliberatori (factori de eliberare release factors) si
hormoni inhibitori care controleaz secretia hormonilor
din adenohipofiz. Transportul factorilor de eliberare si
inhibitie ctre adenohipofiz se face prin intermediul
sistemului port hipotalamico-hipofizar, alctuit din vase
sanguine mici. Odat ajunsi n adenohipofiz, acesti
hormoni stimulatori sau inhibitori actioneaz asupra
celulelor glandulare, reglndu-le secretia. Adeno-
hipofiza secret ase hormoni importanti care sunt
sintetizati n tabelul 3.2, alturi de factorii de eliberare
si de inhibitie.
Neuronii neuroendocrini magnocelulari, locali-
zati n nucleul supraoptic si n cel paraventricular (fig.
3.22), elibereaz oxitocin si vasopresin (hormonul
antidiuretic ADH). Neurohipofiza, spre deosebire de
adenohipofiz, nu secret nici un hormon. Ea este
alctuit din pituicite si are un rol structural, de
sustinere, a unui numr important de fibre si tracturi
care si au originea n hipotalamus.
Curs de neuropsihologie 39
Hormoni
inhibitori
Hormoni de
eliberare
Celule
glandulare
Hormon Aciuni fiziologice
ai hormonului
de crestere
(GIH sau
GHRIH sau
somatostatin)
a hormonului de
crestere (GHRH)
Somatotrope
Hormon de crestere
(GH
somatotropin)
Stimuleaz cresterea;
stimuleaz lipoliza; inhib
actiunile insulinei asupra
metabolismului lipidic si
glucidic
-
a corticotropinei
(CRH)
Corticotrope
Hormon
adrenocorticotrop
(ACTH
corticotropin)
Stimuleaz sinteza de
glucocorticoizi si androgeni
corticosuprarenalieni;
mentine dimensiunile zonei
fasciculate si reticulare a
corticosuprarenalei
-
a tirotropinei
(TRH)
Tirotrope
Hormon stimulator
al secretiei
tiroidiene (TSH
tirotropin)
Stimuleaz sinteza de
hormoni tiroidieni; mentine
dimensiunile celulelor
foliculare
-
a gonadotropinelor
(GnRH)
Gonadotrope
Hormon foliculo-
stimulant (FSH)
Stimuleaz dezvoltarea
foliculilor ovarieni; regleaz
spermatogeneza testicular
Hormon luteinizant
(LH)
Induce ovulatia si formarea
corpului galben ovarian;
stimuleaz productia
ovarian de estrogeni si
progestogeron; stimuleaz
sinteza testicular de
testosteron
ai prolactinei
(PIH)
a prolactinei (PRH)
Lactotrope,
mamotrope
Prolactin (PRL)
Stimuleaz secretia si
sinteza lactat
Tabelul 3.2. Celulele i hormonii adenohipofizei, aciunea lor fiziologic i factorii lor de eliberare i de inhibiie,
secretai de nucleii hipotalamici. Abrevierile sunt pentru termenii din limba englez, acestea fiind standard.
Fig. 3.22. Anatomia funcional a hipofizei.
Curs de neuropsihologie 40
Prin urmare, controlul SE se realizez n dou
moduri : n mod direct (prin secretia produsilor
neuroendocrini n circulatia general cu ajutorul vaselor
sangvine ce strbat neurohipofiza) ; n mod indirect
(prin deversarea factorilor de eliberare sau inhibitie n
sistemul port hipotalamico-hipofizar, de unde ajung n
adenohipofiz, ale crei celule, vor elibera hormoni n
circulatia general).
Printr-un mecanism de tip feed-back,
hipotalamusul si hipofiza sunt influentate de hormonii
secretati de organele tint, iar hipotalamusul poate fi
influentat si de hormonii hipofizari prin tractul tubero-
infundibular si sistemul port.
Hipotalamusul are aferente si eferente neurale
(fascicule nervoase) si umorale (hormoni). Astfel,
neuronii hipotalamici particip la patru clase de reflexe:
1) reflexe conventionale (input si output neurale), 2)
reflexe n care inputul este neural i outputul este
umoral, 3) reflexe n care inputul este umoral i
outputul este neural si 4) reflexe n care inputul i
outputul sunt umorale. Un comportament tipic implic
mai mult de unul din aceste reflexe. De exemplu, emo-
tia are un input senzorial (neural) si ouputuri motorii
viscerale (neurale) si endocrine (hormonale).
n afara hormonilor reglatori (eliberatori si inhi-
bitori) ai hipotalamusului, se mai gsesc si alte peptide :
opioide, betaendorfina, enkefalina, angiotensina II,
substanta P, neurotensina etc. (vezi neurotransmittorii,
cap. 2.4.). Pentru unele din aceste peptide se cunoaste
structura, dar nu se cunoaste efectul fiziologic.
Infundibulul contine o concentratie mare de
acetilcolin (ACh), iar sistemul tubero-infundibular
contine dopamin (DA) care nu actioneaz asupra
hipofizei.
2. Controlul SNV. Hipotalamusul este cel mai
important centru subcortical de reglare a activittii
simpatice si parasimpatice. Aceast functie asigur
mentinerea unei stri interne adecvate supravietuirii
organismului.
Controlul activittii simpatice este legat de
zona lateral si dezona posterioar. Stimularea acestor
regiuni activeaz partea toracico-lombar, intensificnd
activittile somatice si metabolice caracteristice stresu-
lui emotional, atacului si fugii. Se observ reactii ca :
dilatarea pupilelor, piloerectie, tahicardie, cresterea
tensiunii arteriale etc. Distrugerea hipotalamusului
posterior produce letargie, somn anormal si scderea
temperaturii datorat diminurii generale a activittii
somatice viscerale.
Controlul activittii parasimpatice se afl n
relatie direct cu zona anterioar si cu zona medial a
hipotalamusului. Stimularea acestor regiuni duce la
cresterea rspunsurilor autonome sacrate si vagale (ale
nervului vag) caracterizate prin bradicardie,
vasodilatatie periferic, cresterea acidului gastric, a
tonusului tractului digestiv si vezical si, ocazional,
scderea tensiunii arteriale si mioz. Cushing a observat
c excitarea hipotalamusului anterior provoac o
hipermotiliate a tractului gastrointestinal si o grbire a
evacurii. De asemenea, a obtinut experimental
tulburri ale stomacului si duodenului, ulcere acute si
perforarea acestora dup lezarea hipotalamusului.
Hipotalamusul este implicat n numeroase
afectiuni cardiovasculare, respiratorii, gastrointestinale
si n epilepsia diencefalic care vor fi discutate ceva
mai jos, n cadrul sindromului hipotalamic.
3. Controlul sistemului motivaional. Strile
motivationale reprezint impulsuri sau stimulente
determinate de anumite conditii interne, care mping
animalele si omul la actiune sau directioneaz
comportamentul voluntar n vederea satisfacerii unor
necesitti corporale. Implic mecanisme si comporta-
mente complexe, a cror intensitate si directionalitate
are drept scop reglarea temperaturii corporale,
satisfacerea foamei, setei i cerintelor sexuale. n
principiu, strile motivationale sunt mai putin corelate
cu factorii externi si mai degrab cu homeostazia
intern (desigur, este vorba de motivatia fiziologic,
care nu include trebuinte de securitate, de dragoste,
aspiratii, convingeri s.a.m.d.). Vrsta are un rol
important, deoarece la bebelusi functia homeostatic nu
este reglat dect partial din interior, restul fiind pus pe
seama adultului, iar la btrni apare degradarea.
Hipotalamusul exercit un mecanism de control
de tip servomecanism care controleaz mrimile. Acest
control fiziologic de reglare a variabilelor homeostatice
permite organizarea gndirii noastre si cu privire la
unele sisteme mai evoluate, complexe. Mecanismul
functioneaz prin msurarea valorilor anumitor
parametri si compararea lor cu o valoare etalon. Dac
cele dou valori sunt egale, hipotalamusul nu intervine,
iar dac nu sunt egale, apar senzatiile specifice sau sunt
rezolvate direct de ctre hipotalamus (de ex, n cazul
foamei, glicemia sczut semnaleaz o dereglare sau o
stimulare exterioar : vederea sau mirosirea hranei).
Reglarea temperaturii corpului este realizat
exclusiv de ctre hipotalamus. Partea anterioar este
centrul implicat n risipirea cldurii, iar cea
posterioar n producerea cldurii. Dac este lezat
Curs de neuropsihologie 41
partea anterioar se ajunge la hipertermie cronic, iar
dac este lezat partea posterioar se ajunge le
hipotermie rapid n conditii vitrege de mediu. Dac
leziunea se ntinde si asupra altor regiuni, cum ar fi
corpii mamilari, simptomele se pot agrava. Pentru
reglarea temperaturii este necesar integrarea a cel putin
patru procese asociate cu functiile endocrine si vegetati-
ve ale hipotalamusul : 1) sesizarea cresterii sau scderii
temperaturii din snge de ctre neuronii sensibili la
temperatur (termoreceptori) ; 2) elibrarea TRH de
ctre hipotalamus, care conduce la secretia de TSH a
adenohipofizei (tabelul 3.2), rezultnd n secretia
hormonilor tiroidieni pentrucreterea ratei metabolice ;
3) dilatarea sau constrictia vaselor sanguine care
rezult n risipirea si, respectiv, conservarea cldurii ;
4) activarea unor comportamente motorii ca gfiala
(pentru risipirea cldurii) sau tremuratul (pentru
conservarea cldurii).
Reglarea foamei i a aportului de hran se afl
sub controlul a doi centri hipotalamici : 1) nucleul
ventromedial i tesutul nconjurtor (centrul satiettii ;
lezarea produce hiperfagie si obezitate) si 2) grupul
nuclear lateral (centrul hrnirii ; lezarea bilateral
produce afagie i moarte, chiar n conditiile hrnirii
fortate !).
Reglarea setei
1
este legat de eliberarea ADH
(antidiuretic hormone / vasopresina) de ctre nucleii
supraoptic si paraventricular n circuitul sanguin de la
nivelul neurohipofizei. S-a demonstrat c stimularea
nucleului paraventricular activeaz un mecanism care
induce retentia apei din rinichi. Apa din organism este
stocat n celule si n afara celulelor (n snge si alte
fluide). Evacuarea apei se face fie cu ajutorul rinichilor
(urina), fie cu ajutorul glandelor sudoripare, fie sub
form de vapori, din plmni si n toate cazurile vine
din circuitul sanguin. Receptorii din presiunii din ri-
nichi, inim si vasele de snge mari detecteaz pierde-
rea de ap si activeaz neuronii senzoriali care transmit
un semnal celor din hipotalamus, determinndu-i s
elibereze ADH. Actiunea AHD-ului face ca rinichii s
pstreze apa, n loc s o transforme n urin. De aseme-
nea, rinichii sunt impulsionati s eliberezerenin, care
prin reactie chimic cu o substant din snge, produce
angiotensina, hormon care activeaz neuronii din struc-
turile profunde ale creierului, crend senzatie de sete.
Aceasta este setea extracelular. Pe lng ea, mai
exist si setea intracelular, care este determint de
dishidratarea celular provocat de concentratiile cres-
cute de sare. Cnd bem, nlocuim apa din snge, redu-
1
Nepredat.
cnd concentratia de sare, ceea ce permite apei s
revin n neuroni si alte celule. Acesta este motivul
pentru care consumul srii provoac sete.
Reproducere i functiile sexuale. Neuronii din
nucleul preoptic si ventromedial sunt sensibili la
estrogen si androgen si declanseaz productia de hor-
moni corespunztori (GnRH) ce controleaz productia
si eliberarea de gonadotropine din hipofiza anterioar.
Studiile experimentale au evidentiat diverse functii ale
altor nuclei. Spre exemplu, stimularea corpilor mamilari
la veverit provoac erectia penisului.
Alte funcii ale hipotalamusului
2
. Hipotala-
musul este implicat si n alte functii foarte importante,
cum ar fi reglarea metabolismului (glucidic, lipidic,
protidic, ureic), reglarea comportamentului afectiv, re-
glarea memoriei, reglarea ritmului circadian, somnului
s.a. S analizm succint cteva dintre acestea.
Reglarea comportamentului afectiv. Am artat
mai sus c hipotalamusul are un rol n reglarea cardio-
vascular si a altor functii autonomice (vegetative) ale
SN. Aceste functii includ componente ale diferitelor
forme de comportament emotional, care sunt integrate
n diferite regiuni hipotalamice. Mai multe forme de
comportament emotional au fost identificate, n special
datorit experimentelor pe pisici.
Stimularea prtii mediale a hipotalamusului
pisicii provoac o manifestare de furie defensiv.
Aceasta se manifest prin piloerectie, scuipat, mrit,
retragerea urechilor pe spate, lovirea obiectelor mobile
din jur. Reactia este defensiv n esent si apare, n mod
natural, cnd pisica se simte n pericol. n afar de hipo-
talamus, n reactia de furie defensiv sunt implicate si
alte structuri cerebrale, cum ar fi amigdala, hipocampul,
aria septal si cortexul prefrontal.
Stimularea prtii laterale a hipotalamusului
provoac o form diferit de comportament agresiv, un
comportament de tipul atacului prdtorilor. Pisica, ce
n mod normal nu atac un soarece, n urma stimulrii,
va pndi si l va ataca pe aceasta, muscndu-l de partea
dorsal a gtului. Aceast form de agresivitatea este
asemntoarea cu cea natural a pisicilor cnd pndesc
un soarece, pe care o stim cu totii. Atacul de prdtor
este influentat si de conexiunile (directe sau indirecte)
cu sistemul limbic, precum si de actiunea monoamine-
lor (vezi cap. 2.4. si tabelul 2.1).
Partea lateral si partea medial a hipotalamu-
sului se inhib una pe cealalt, sub aspectul acestor
forme de agresivitate. Cu alte cuvinte, cnd animalul
2
Nepredate.
Curs de neuropsihologie 42
este n stare de furie defensiv, se va excita puternic
zona medial, iar cea lateral va fi n stare de repaus si
invers pentru atacul de prdtor.
Reactia de fug, scpare (fleeing) este provoca-
t prin stimularea mai multor prti ale hipotalamusului.
Pisica ncearc disperat s ias din cusc. Aceast
reactie este influentat de fibrele ascendente provenite
de la substanta cenusie periapeductal din mezencefal.
Apatia este asociat cu lezarea tracturilor
simpatice ale hipotalamusului, iar euforia este uneori
considerat ca fiind provocat de hipotalamus.
Ritmul circadian este un ciclu de comporta-
mente sau modificri fiziologice care dureaz aproxi-
mativ 24 de ore. Activitatea ritmului circadian poate fi
divizat n dou faze : faza activ si faza inactiv
(odihn, somn). Relatiile dintre cele dou faze difer
dup conditiile de antrenament, putnd fi scurtate sau
prelungite. Nucleul suprachiasmatic (legat de cel supra-
optic si n partea anterioar) primeste impulsuri retinale
(de la ochi) care par s fie foarte importante pentru
controlul ritmurilor circadiene a unor hormoni sexuali,
corticosteron si melatonin (vezi cap. 3.4.3. pt. melat.).
Aceste ritmuri hormonale sunt asociate cu ritmurile
circadiene pentru temperatura corpului si comportamen-
tul sexual. Este posibil ca aria preoptic s joace un rol
important n ritmul circadian al eliberrilor de LH (leu-
teinizant hormone) din adenohipofiz, deoarece neuro-
nii GnRH (gonadotrope realeasing hormone) sunt loca-
lizati n aceast regiune.
Somnul este controlat, n principal, de substan-
ta reticulat si de hipotalamus. Experimentele fcute pe
sobolani au artat c lezarea hipotalamusului posterior
produce perioade prelungite de somn. Cercetri ulte-
rioare au artat c somnul prelungit a fost produs mai
degrab datorit ntreruperii tracturilor care vin de la
formatiunea reticulat dect lezrii nucleilor hipotala-
mici. Studii recente sugereaz c aria preoptic si subs-
tantia innominata functioneaz mpreun cu alte regiuni
colinergice si monaminergice din formatiunea reticula-
t, pentru a regla ciclul somn/veghe.
Patologia hipotalamusului. Distrugerea tesu-
tului nervos hipotalamic poate provoca diverse tulburri
ale functiilor prezentate mai sus. Tulburrile hipotala-
musului pot rezulta din leziuni specifice cauzate de
tulburri vasculare, inflamatia tesutului hipotalamic sau
tumori, care, de regul, apar pe planseul ventriculului
III sau pe glanda pituitar.
Tulburrile sistemului endocrin sunt consecinta
absentei hormonilor reglatori hipotalamici (tabelul 3.2)
ce influenteaz adenohipofiza. Lezarea hipotalamusului
sau a sistemului port hipofizar conduce la reducerea se-
cretiei tuturor hormonilor adenohipofizei, cu exceptia
PRL. Prin urmare, simptomele ntlnite vor fi : hiposu-
prarenalism (legat de ACTH), hipotiroidism (TSH) si
anomalii ale ciclului sistemului reproductor (GnRH).
Dezvoltarea sexual este tulburat n cazul
anumitor leziuni ale hipotalamusului. Cele mai
cunoscute simptome sunt hipogonadismul si pubertatea
precoce.
Pubertatea precoce se refer la spermatogeneza
si secretia prea timpurie de androgen, la bieti si apari-
tia menstruatiei si secretiei prea timpurii a estrogenului,
la fete.
Hipogonadismul, dup cum i este si numele,
reprezint un deficit de secretii de gonade, care conduce
la ntrzierea dezvoltrii caracteristicilor sexuale secun-
dare (la fete subdezvoltarea snilor, lipsa menstruatiei
si a prului pubian ; la bieti subdezvoltarea organelor
genitale, lips pilozittii faciale, toracice si pubiene).
De obicei, hipogonadismul este asociat cu obezitate,
somnolent si diabet insipid. Dac apare o traum me-
canic unilateral, tulburarea sexual este usoar si trece
repede (este recuperat de cealalt parte). Prin urmare,
doar n cazul leziunilor bilaterale exist tulburri inten-
se legate de dezvoltarea functiilor sexuale.
Tulburrile legate de control SNV includ tulbu-
rri cardiovasculare, tulburri respiratorii, tulburri
gastro-intestinale si epilepsia diencefalic.
Tulburrile cardiovasculare sunt corelate dese-
ori cu afectiunile hipotalamusului si ale trunchiului
cerebral. Din acestea, amintim hipertensiunea, variate
tipuri de aritmii cardiace, modificri EKG, din care
unele pot simula un infarct miocardic acut, la bolnavii
Ir antecedente cardiace. Hipertensiunea pare s fie
provocat de secretia excesiv de ACTH (adrenocorti-
cotrope hormone), care, la rndul ei, este o urmare a
hipersecretiei de CRH (corticotropine releasing hor-
mone) din nucleii hipotalamici.
Tulburrile respiratorii sunt foarte frecvente n
cadrul sindromului hipotalamic. Aceasta se ntmpl
deoarece hipotalamusul anterior contine neuroni care
comand ciclul si ritmul respirator. Cele mai frecvente
afectiuni sunt edemul pulmonar (acumularea apei n
plmni) si hemoragia pulmonar.
Tulburrile gastro-intestinale produse de ctre
hipotalamus includ, n principal, hemoragiile gastrice si
ulceratiile. Experimentele pe animale au artat c leziu-
Curs de neuropsihologie 43
nile nucleilor tuberali provoac eroziuni superficiale
sau ulceratii ale mucoasei gastrice, n absenta hiperaci-
dittii (ulcere Cushing) care este conditia normal a
ulcerului. Leziuni gastrice asemntoare au fost semna-
late si la pacientii cu diverse leziuni intracraniene.
Epilepsia diencefalic a fost descris pentru
prima oar de ctre Penfield (1929), la un pacient care
prezenta dureri de cap si dou tipuri de crize. n primul
tip de criz pacientul acuza ameteal si avea tendinta de
a cdea. n cel de-al doilea tip, fata si bratele se nro-
seau, respiratia ncetinea, transpira, i curgea saliv din
gur si i ieseau ochii n afar (exoftalmie). Diagnosti-
cul de epilepsie diencefalic este cel mai bine justificat
la pacientii care au o disfunctie hipotalamic precis
identificat.
Tulburri ale controlului termic. Febra hipota-
lamic se manifest prin cresterea neregulat a
temperaturii corpului, lipsa transpiratiei, temperatur
central mai ridicat dect temperatura periferic.
Diagnosticarea se face prin eliminarea altor posibile
afectiuni.
Sindromul termic hipotalamic apare n conditii
legate de diferite afectiuni ale hipotalamusului si este
prezent n 5 forme: poikilotermie, hipertermie sustinut,
hipertermie paroxist, hipotermie sustint, hipotermie
paroxist.
Poikilotermia reprezint incapacitatea mentinerii
temperaturii centrale la un nivel constant,
indiferent de temperatura ambiental. Fluctuatia
trebuie s fie mai mare de 2C pentru a putea
vorbi de poikilotermie. Rezult din lezarea sau
disfunctia centrilor integratori si efectori ai
termoreglrii din hipotalamusul posterior si
mezencefalul rostral. Unii pacienti nu sunt surpinsi
de conditia lor si nu arat nici un semn de
disconfort sau activitate reglatorie la stresul termic.
Pentru a fi produs o poikilotermie, lezarea
trebuie s fie bilateral.
Hipertermia sustinut reprezint perturbarea
mecanismelor de pierdere a cldurii si stimularea
mecanismelor de producere a ei. Apare prin lezarea
unor nuclei din hipotalamusul anterior si const n
cresterea necontrolat a temperaturii corpului ca
urmare a producerii de cldur.
Hipertermia paroxist are un caracter episodic si
se manifest prin frisoane cu nalte crosete febrile
(pusee de crestere a temperaturii) si alte fenomene
vegetative.
Hipotermia sustinut apare fie prin lezarea
mecanismelor de producere a cldurii, fie prin
stabilirea unui centru anormal inferior. Hipotermia
sustinut se produce prin lezarea hipotalamusului
anterior si apare foarte rar n patologie, mai
frecvent fiind :
Hipotermia paroxist care const n episoade de
scdere a temperaturii corpului, cu o frecvent
variabil n functie de zi. Debuteaz brusc prin
transpiratie, nrosirea pielii, scderea temperaturii
corpului pn la 32C. Dureaz de la cteva minute
la cteva zile. Simptome asociate : oboseal,
diminuarea activittii cerebrale, hipoventilatie,
hipotensiune, aritmie cardiac, ataxie, lcrimare.
Mecanismele de producere i eliberare a cldurii
functioneaz normal, dar pentru valoarea de 32C!
Hipotermiile paroxiste pot fi : usoare (35-32C),
moderate (32-24C), grave (sub 24C). Hipotermia
paroxist grav este ireversibil, provoac com
i deces prin fibrilatie ventricular.
Tulburri ale foamei i aportului de hran.
Comportamentul la foame este legat n mare msur de
mentinerea greuttii corporale la o valoare relativ fix.
n conditiile variatiei aportului alimentar apar mecanis-
me ce aduc greutatea la nivelul normal (normalul
organismului).
Experimental, s-a demonstrat c lezarea
bilateral a nucleilor ventromediali duce la obezitate,
hiperfagie, comportament slbatic.
n general, anorexia este considerat o boal
psihic, dar poate aprea si ca urmare a dereglrii
functiilor hipotalamice. Este nsotit de scderea n
greutate, de amenoree (absenta fiziologic a menstrua-
tiei) si de alte tulburri endocrine (de exemplu,
poikilotermie).
Lezarea hipotalamusului lateral produce afagie
i emaciere. Emacierea reprezint lipsa foamei urmat
de scderea n greutate. Este asociat cu miscri oculare
nistagmoide, comportament afectiv inadecvat, apatie,
iritabilitate (la copii) sau, dimpotriv, hiperactivitate n
pofida slbirii. n putinele cazuri de supravietuire, peste
2 ani, a revenit apetitul, emacierea a cedat astfel nct
copii au devenit obezi hipotalamici, iar iritabilitatea si
chiar furia au tins s ia locul euforiei si hiperactivittii.
Hipotalamusul nu are rol numai n reglarea
cantittii de hran, ci si n reglarea calittii acesteia prin
aportul de lipide, glucide si proteine. Pn la vrsta de
un an, hipotalamusul nu controleaz aportul de lipide.
Sindromul lipidic apare n cazul lezrii bazei
hipotalamice si prezint trei variante :
Curs de neuropsihologie 44
1. Sindromul Babinski-Frhlich : obezitate si infantil-
ism genital ; este provocat de lezarea hipotalamu-
sului bazal si medial ;
2. Sindromul Lawrence-Moon-Bardet-Biedl : obezi-
tate, hipogonadism, deficient mintal, deformatii
craniene si dentare, malformatii congenitale
(polidactilie) ; pare a avea caracter ereditar si nu
prezint mereu leziune hipotalamic vizibil ;
3. Sindromul Prader-Labhart-Willi : obezitate, hipo-
gonadism, statur mic, predispozitie pentru diabet
zaharat ; cel mai probabil, nu este ereditar.
Diabetul insipid (DI), o tulburare a echilibrului
hidric (nepredat), survine ca urmare a absentei
vasopresinei (ADH), produs de nucleul supraoptic si
cel paraventricular si eliberat direct n circuitul
sangvin, la nivelul neurohipofizei (hipofiza anterioar).
n absenta ADH, apa nu este reabsorbit de ctre
rinichi, iar productia urinar este extrem de ridicat. DI
poate fi temporar, cnd este dat de o leziune intraselar
sau a infundibulumului, sau permanent, cnd este dat de
lezarea eminentei mediale a tuber cinereum (fig. 3.22)
sau a hipotalamusului anterior. Cauzele cele mai
frecvente sunt cele amintite pentru ntreg sindromul
hipotalamic.
Tulburrile somnului i tulburrile afective sunt
prezentate n cadrul fiziologiei hipotalamusului (vezi
mai sus, alte functii ale hipotalamusului).
3.5. TELENCEFALUL
Telecenfalul reprezint, la om, cel mai proeminent si
cel mai evoluat segement al sistemului nervos central
(SNC). La vrsta de 6 ani, atinge greutatea creierului
adultului: la brbati aproximativ 1.4 Kg, iar la femei
aproximativ 1.25 Kg. Se mai numeste si creierul mare
sau cerebrum. Se compune din emisferele cerebrale,
comisurile si cavittile lor.
Emisferele cerebrale reprezint cea mai volu-
minoas mas nervoas a ntregului SN. Ele sunt sepa-
rate de ctre fisura cerebral longitudinal sau fisura
interemisferic, si sunt conectate prin intermediul cor-
pului calos. Sunt alctuite din: sistemul limbic,
ganglionii bazali, substanta alb la interior si scoarta
cerebral, alctuit din substant cenusie, la exterior.
Fiecare emisfer are trei fete: extern sau
superolateral (fig. 3.25), intern sau medial (fig.
3.26) si inferioar saubazal (fig. 3.27). De asemenea,
fiecare emisfer are si cte trei margini: superioar
(superomedial), inferolateral si inferomedial (ntre
suprafata intern si cea medial).
La extremitatea anterioar a emisferei se afl
polul frontal, la cea posterioar polul occipital, iar
extremitatea anterioar a lobului temporal formeaz
polul temporal (fig. 3.25, 3.26).
Suprafaa extern (fig. 3.25) sau lateral este
adaptat convexittii cutiei craniene si este brzdat de
numeroase cirumvolutii (girusuri) separate ntre ele de
anturi (scizuri sau sulcusuri). Aria cortexului cerebral
uman este de aproximativ 2200 cm
2
. O treime din
aceast arie este vizibil la suprafat, restul fiind ascuns
n sciziuri si fisuri. Cele trei santuri majore, care mpart
emisferele cerebrale n patru lobi cerebrali (lobul
frontal, lobul temporal, lobul parietal i lobul occipital,
fig. 3.23) sunt:
- Scizura lateral (scizura lui Sylvius);
- Scizura central (scizura lui Rolando);
- Scizura parieto-occipital (scizura perpendicu-
lar extern).
Lobul frontal reprezint portiunea cea mai ante-
rioar a emisferelor cerebrale, desprtit de lobul parie-
tal de ctre scizura lui Rolando si de lobul temporal de
ctre scizura lui Sylvius. Imediat n partea anterioar a
santului central se afl girusul precentral, o important
arie motorie. De aici pleac att tracturile piramidale,
ct si cele extrapiramidale. n partea anterioar, girusul
precentral este delimitat deantul precentral. Functiile
Fig. 3.23. Lobii cerebrali.
Curs de neuropsihologie 45
lobului frontal includ initierea miscrilor voluntare,
analiza superioar a datelor senzoriale si provocarea
rspunsurilor specifice personalittii. De asemenea,
mediaz rspunsuri legate de memorie, afectivitate,
ratiune, judecat, planificare si comunicare verbal
(centrul inteligentei umane). ntorcndu-ne la povestea
lui Phineas Gage de la nceputul cap. 1, putem observa
cauza fiziologic a tragicei sale schimbri. De
asemenea, ne putem da seama c dac lobul frontal ar fi
continut centri vitali, Phineas ar fi murit pe loc.
Lobul parietal este asezat posterior de scizura
lui Rolando si se ntinde pn la scizura parieto-occipi-
tal. n partea inferioar, este delimit de (ramul poste-
rior al) santul(ui) lateral si o prelungire posterioar ima-
ginar a acestuia (fig. 3.23 n comparatia cu fig. 3.25),
prin care este unit de scizura parieto-occipital. Imediat
n partea posterioar a scizurii lui Rolando se afl
girusul postcentral, o important arie senzorial.
Aceasta rspunde la senzatiile cutanate si proprioceptiv-
kinestezice de pe suprafata ntregului corp. mpreun cu
girusul precentral, formeaz aria functional primar
(senzorio-motorie).
Lobul temporal este situat sub lobul parietal si,
partial, sub portiunea posterioar a lobului anterior. Este
delimitat superior de scizura lui Sylvius si de aceeasi
prelungire imaginar a acesteia care delimiteaz si lobul
parietal (fig. 3.23 n comparatia cu fig. 3.25). Suprafata
lateral este mprtit de dou santuri n trei
circumvolutiuni paralele (girusul temporal superior,
mijlociu si inferior). Functia principal a lobului
temporal este perceptia semnalelor sonore. Cortexul
auditiv primar pare a fi localizat n girusurile lui
Henschl, din girusul temporal superior (neexistente n
fig. 3.25).
Lobul occipital se afl cel mai posterior,
napoia liniei arbitrare care uneste santul parieto-
occipital cu incizura preoccipital (fig. 3.23, 3.25).
Dedesuptul lobului occipital se afl cerebelul, de care
este separat prin tentorium cerebelli (cortul
cerebelului). Principala functie a lobului occipital tine
deperceptia vizual.
Insula (lui Reil) este un lob care nu se observ
la suprafata emisferelor, fiind situat n profunzimea
santului lateral (fig. 3.24). Suprafata insulei este
divizat, de ctre santul central al insulei, ntr-o
portiune anterioar (mai mare) si una posterioar (mai
mic). Partea anterioar este mprtit de santuri putin
adnci n trei sau patru girusuri (circumvolutii) scurte,
iar partea posterioar este constituit dintr-un singur
girus lung, divizat adesea la captul su posterior.
Insula a rmas fix de-a lungul evolutiei
filogenetice, fiind acoperit de neocortexul
nconjurtor, pe msur ce acesta din urm s-a
dezvoltat. La om, este voluminoas si se dezvolt din
cortexul olfactiv. Se stiu putine desprefunctiile sale, n
afar de faptul c integreaz unele activitti cerebrale.
Se pare c are un rol si n memorie.
Suprafaa medial este vizibil printr-o sectiune
mediosagital a creierului, care desparte cele dou
emisfere (fig. 3.26). Se poate observacorpul calos, cea
mai mare formatiune de substant alb a emisferelor
cerebrale, antul cingulat, n partea anterioar (mparte
zona respectiv n exterioar si inferioar), girusul
cingulat, sub santul cingulat, fornixul, sub corpul calos
etc. Regiunea posterioar este traversat de dou san-
turi : scizura parieto-occipital care desparte lobul
occipital de lobul parietal si scizura calcarin a lobului
occipital.
Suprafaa inferioar (fig.3.27) este delimitat de
scizura lui Sylvius ntr-o parte anterioar, mai mic si
una posterioar, mai mare, fiecare zon avnd functii
distincte.
npartea anterioar, paralel de fisura cerebral
cerebral, se afl antul olfactiv, acoperit de bulbul si
tractul olfactiv (n fig. 3.27 se poate vedea bulbul si
tractul n partea dreapt si santul n partea stng).
Medial de aceste formatiuni se afl girus rectus. Restul
suprafetei este mprtit de un sant n form de H
(orbital) n patru girusuri orbitale (anterior, posterior,
lateral si medial).
Partea posterioar este traversat de ctre
antul colateral si antul occipitotemporal. Santul rinal
are directia celui colateral si separ lobul temporal de
uncus. n partea posteromedial se poate observaistmul
girusului cingulat (vezi suprafata medial), iar posterior
de acesta spleniul corpului calos. De asemenea, se
mai poate observagirusul inferior al lobului temporal si
girusul parahipocampic.
Fig. 3.24. Insula (lui Reil).
Curs de neuropsihologie 46
Fig. 3.25. Suprafaa lateral (extern) i cea medial a emisferelor cerebrale. n text, pentru majoritatea girusurilor este
folosit termenul echivalent de anuri (detaliu). Din motive de spaiu, suprafaa inferioar a fost inclus n figura urmtoare.
Curs de neuropsihologie 47
Fig. 3.26. Viziune inferioar asupra creierului ce relev suprafaa inferioar a emisferelor cerebrale i alte
structuri anatomice.
Curs de neuropsihologie 48
3.5.1. Sistemul (lobul) limbic
1
Prtile vechi din punct de vedere filogenetic ale
telencefalului, mpreun cu zonele sale marginale si
conexiunile sale cu centri subcorticali, poart denumi-
rea colectiv de sistem limbic sau lob limbic (limb
margine). Structurile anatomice ale sistemului limbic
nconjoar diencefalul (fig. 3.27-A) si unele fac parte
din acesta (hipotalamusul, de exemplu). Spre deosebire
de structurile parcurse pn acum, sistemul limbic nu
este o structur organizat topic, ci mai degrab un grup
de nuclei si arii corticale cu functii asemntoare. Se
pot deduce aceste functii, dac precizm c a mai fost
numit creierul emotional sauvisceral (termeni introdusi
de MacLean, 1958). Structurile anatomice care compun
sistemul limbic sunt incerte, vag si divers definite.
Broca a folosit pentru prima dat termenul de
lob limbic (1878), pentru a caracteriza structurile primi-
tive ale telencefalului ce formeaz un inel n jurul trun-
chiului cerebral. Multe structuri ale sistemului limbic au
form arcuit si sunt situate ntre diencefal si cortexul
cerebral. Celebrul circuit emotional al lui Papez (1937),
care includea si sistemul limbic, a trezit interesul asupra
acestuia din urm.
Sistemul limbic include hipotalamusul si un
grup de structuri interconectate ale telencefalului
(girusul cingulat, parahipocampic, subcalosal, septul,
aria paraolfactiv, nucleul talamic anterior, portiuni
ale ganglioni bazali, hipocampul i amigdala
majoritatea se pot observa n fig. 3.25 si 3.26). n jurul
ariilor subcorticale ale sistemului limbic este situat
cortexul limbic, reprezentat de fiecare parte a creierului
de ctre un inel cortical (aceast structur este arbitrar
si sintetic, n literatura de specialitate ntlnindu-se
multe alte formatiuni, precum si mai putine ; pentru
maximul de structuri anatomice ce au fost incluse n
cadrul sistemului limbic, vezi Tratatul de neuropsiholo-
gie, vol. 1, Dnil, Golu, 2006, p. 364).
Animalele care au un sistem limbic slab dezvol-
tat (pestii, reptilele etc.) desfsoar activittile de
hrnire, atac, fug si mperechere (cei patru F, vezi
hipotalamusul) prin intermediul unor comportamente
stereotipe. La mamifere, sistemul limbic pare s inhibe
unele din aceste comportamente instinctive, permitnd
organismului s fie mai flexibil si mai capabil s se
adapteze la mediu. Concret, functiile componentelor
sistemului limbic tin de controlul comportamentului
emotional i motivational, de memorie si de controlul
1
Din aceast unitate nu a fost predat dect amigdala.
SNV. Am analizat pe larg hipotalamusul n cadrul
diencefalului (unitatea 3.4.5.), iar n continuare vom
analiza celelate dou componente importante ale
sistemului limbic, hipocampul si amigdala (fig. 3.27).
1) HIPOCAMPUL
Hipocampul este localizat n profunzimea
lobului temporal, ptrunznd n ventriculul lateral.
Numele de hipocamp se datoreaz asemnrii acestei
structuri cu un clut sau un monstru de mare (gr.
hippokampos), dup cum se poate observa n fig. 3.28.
n general, neuroanatomia foloseste termenul de
formatiune hipocampic, n cadrul creia intr
hipocampul propriu-zis (cornul lui Ammon), girusul
dintat si subiculumul (fig. 3.28, dar pentru o mai bun
ntelegere topografic se pot studia si fig. 3.25 si 3.26).
n partea inferioar, formatiunea hipocampic este aco-
perit de ctre girusul parahipocampic sau cortexul
entorinal (Fig. 3.25, 3.26, 3.28), care se continu cu
Fig. 3.27. (A) Localizarea structurilor sistemului limbic n
cadrul emisferelor cerebrale; (B) Hipocampul i amigdala.
Curs de neuropsihologie 49
subiculumul, care, la rndul lui, se continu cu cornul
lui Ammon. Pe partea dorsal a hipocampului (cornul
lui Ammon) se afl un strat gros de fibre, numit fimbria
hipocampului (fig. 3.27, 3.28). Sub fimbria se afl
girusul dintat (fig. 3.28). La exterior, hipocampul este
acoperit de un strat de fibre numit alveus (fig. 3.28).
Santul hipocampic separ girusul dintat de girusul
parahipocampic (cortexul entorinal).
Hipocampul sau cornul lui Ammon este subdi-
vizat n patru prti, n functie de ntindere, mrimea si
densitatea celulelor. Cele patru prti sunt notate cu CA,
de la cornu ammonis (fig. 3.28) si au forma literei C.
Acesta sunt :
1. Cmpul CA1 : contine celule piramidale mici, ase-
zate cel mai aproape de subiculum ; celulele de aici
prezint un interes special, deoarece sunt predispu-
se la anoxie (cantitate inadecvat de oxigen n
tesut), mai ales n cazul epilepsiei de lob temporal
(se mai numestesectorul lui Sommer) ;
2. Cmpul CA2 : o band lung de celule piramidale
mari, situate ntre CA1 si CA3 ;
3. Cmpul CA3 : o band larg de celule piramidale
situat ntre CA2 si CA4 ;
4. Cmpul CA4 : formeaz zona intern a girusului
dintat (mai nou, se pune problema dac se poate
delimita regiunea CA4 de regiunea CA3).
Celulele hipocampului sunt repartizate si n
straturi. Astfel, de la suprafat spre interior avem : 1)
stratul plexiform si alveus, care contin fibre eferente
(din neuronii piramidali) si aferente (de la cortexul
entorinal) ; 2) stratum oriens, care contine dendritele
bazale ale neuronilor piramidali si celule n form de
cos care au efect inhibitor asupra celulelor piramidale ;
3) stratul piramidal si 4) stratul radiatum si lacunosum-
moleculare, care contin dendritele apicale (din vrf) ale
neuronilor piramidali si aferente de la cortexul
entorinal.
Girusul dinat este o structur cortical cu trei
straturi (fig. 3.28), a crui tip principal de celule sunt
neuronii granulari (acestia alctuind stratul granular).
Axonii celulelor granulare (fibre muschiulare) fac
sinaps cu celulele piramidale din
CA3. n profunzimea stratului
granular se afl un strat polimorf,
alctuit din celule piramidale modifi-
cate, iar extern de stratul granular se
afl stratul molecular, alctuit, n
principal, din axonii fibrelor aferente
hipocampice. Stratul molecular este
situat lng stratul molecular al
hipocampului.
Subiculumul (fig. 3.28) sau
cortexul subicular este o regiune de
comunicare ntre cortexul entorinal
(girusul parahipocampic) si hipocamp.
Diferenta histologic esential dintre
hipocamp si subiculum rezid n
faptul c stratul piramidal din
subiculum este semnificativ mai gros
spre deosebire de cel hipocampic, dup cum se poate
observa n fig. 3.28.
Conexiunile hipocampului. Cea mai impor-
tant aferent a hipocampului este alctuit din fibrele
care provin de la cortexul entorinal. Pe aceast cale, n
hipocamp ajung fibre de la centrii olfactivi primari, de
la corpul amigdaloid si diverse regiunil ale neocorte-
xului. Legturi directe ntre bulbul olfactiv si hipocamp
nu au fost demonstrate.
Hipocampul mai primeste aferente de la giru-
sul cingulat, n principal n subiculum. Prin fornix, se
primesc inputuri de la aria septal. Alte surse aferente
includ cortexul prefrontal, regiunea premamilar si sub-
stanta reticulat. Prin inputurile provenite de la diferite
regiuni, hipocampul poate reactiona ca un releu la
evenimentele ce au loc n neocortex si trunchiul cere-
bral, transmitnd informatii despre aceste evenimente la
hipotalamus, care provoac modificri viscerale sau
emotionale la acestea.
n afar de cteva fibre care prsesc hipo-
campul prin stria logitudinal, majoritatea eferentelor
hipocampice iau calea fornixului. Fornixul are o parte
precomisural si una postcomisular. Fornixul preco-
misular contine fibre care se termin n septum, n aria
preoptic si n hipotalamus. Fornixul postcomisular
contine fibre care se termin n corpii mamilari (predo-
minant n nucleul medial al corpilor mamilari), n nu-
cleii talamici anteriori si n hipotalamus. Unele fibre ale
fornixului se extind pn la substanta cenusie aperiduc-
tal a mezencefalului.
Circuitul lui Papez are ca punct de plecare hi-
pocampul. Impulsurile hipocampice ajung la corpii
mamilari pe calea fornixului. De aici, pleac prin fasci-
culul lui Vicq dAzyr (tractul mamilotalamic) ctre
nucleii anteriori ai talamusului, iar mai departe, este
trimis ctre girusul cingulat. De la girusul cingulat,
impulsul ajunge la cortexul entorinal pe calea cingulu-
mului (mnunchi de fibre cinguloentorinale). De aici,
ajunge napoi n hipocamp. J ames Papez (1883-1958) a
Fig. 3.28. Anatomia intern a formaiunii hipocampice (seciune coronal).
Poriunea albastr reprezint ventriculul lateral.
Curs de neuropsihologie 50
descris acest circuit n 1937 (A proposed mechanism of
emotion), observnd calea pe care o urmeaz virusul
rabiei n creierul unei pisici. Acest circuit este implicat
n emotie, dup cum arat si numele articolului lui
Papez, dar s-a descoperit c are un rol si n stocarea
informatiilor (memorie). O parte din componentele
circuitului Papez se pot observa n fig. 3.29.
Fornixul (fig. 3.19, 3.21, 3.25, 3.27) este alc-
tuit, dup cum am vzut, din fibrele hipocampice afe-
rente si eferente, care se arcuiesc n jurul prtii rostrale,
caudale si superioare ale talamusului (fapt vizibil cel
mai bine n fig. 3.21 si 3.27-A). Este compus dintr-o
parte caudal ltit crura sau crus, o suprafat supe-
rioar compact corpul si o parte care se arcuieste
peste partea rostral a talamusului si trece prin hipota-
lamus, ajungnd la corpii mamilari coloanele. Forni-
xul emisferei drepte comunic cu fornixul emisferei
stngi prin comisura fornixului. Aceste componente se
pot observa cel mai bine n fig. 3.25 si 3.27-B.
Fiziopatologia formaiunii hipocampice.
Functiile formatiunii hipocampice includ anumite forme
de nvtare si memorare, control agresivittii si contro-
lul functiilor vegetative si endocrine.
Memoria i nvarea au fost asociate cu func-
tionarea formatiunii hipocampice n urma studiile reali-
zate att pe animale, ct si pe oameni (clinica neurolo-
gic).
n cazul animalelor, s-a descoperit c hipo-
campul genereaz un ritm teta (vezi cap. 4.1.) cnd
animalul se apropie de o tint sau n diferitele faze ale
conditionrii clasice sau operante. Alte studii au artat
c animalele cu leziuni hipocampice continu s ofere
acelasi rspuns ntr-o situatie de nvtare, chiar dac
acesta nu este cel bun. De asemenea, animalul si pierde
si capacitatea de a amna rspunsul.
La om, se pare c, mai ales n primele stadii
ale dezvoltrii creierului uman, hipocampul are un rol
important n luarea deciziilor, prin stabilirea importantei
impulsurilor senzoriale aferente (acesta fiind stimulat
de aproape orice senzatie a subiectului). Prin urmare,
dac hipocampul stabileste c un anumit semnal aferent
este important, atunci informatia respectiv va fi retinu-
t si memorat. Astfel, o persoan se obisnuieste rapid
cu stimulii indiferenti si retine orice experient senzo-
rial care induce senzatia de plcere sau durere (condi-
tionare operant).
Hipocampul este implicat n toate aspectele
memoriei declarative, cum ar fi memoria semantic
(concepte), memoria episodic (ntmplri si succesiu-
nea lor cronologic) si memoria spatial (recunoasterea
localizrii spatiale). Se crede c hipocampul este
responsabil pentru transmiterea informatiilor din
memoria de scurt durat n memoria de lung durat.
n orice caz, n absenta hipocampului, consolidarea pe
termen lung a informatiilor retinute (verbale, simbolice
sau spatiale) este redus sau irealizabil.
Sindromul amnezic (Korsakoff) este una din
cele mai timpurii forme de tulburri ale memoriei care
implic formatiunea hipocampic. Starea de amnezie
(dup cum a fost definit original de ctre Th. Ribot),
include dou mari componente distincte : 1) amnezia
anterograd (diminuarea capacittii sau imposibilitatea
persoanei de a-si aminti informatii stabilite bine n
memorie naintea leziunii, cum ar fi numele, vrsta
s.a.m.d.) si 2) amnezia retrograd (diminuarea capa-
cittii sau imposibilitatea de a pstra, de a nvta lucruri
noi). Aceste dou componente nu au mereu aceeasi
intensitate, una n comparatie cu alta. Se mai vorbeste,
uneori, si de o a treia component confabularea
(pacientul nu-si aduce aminte un lucru, dar si foloseste
imaginatia pentru a da un rspuns). n cadrul sindromu-
lui Korsakoff sunt afectate si alte functii cognitive care
depind de memorie, dar nu la fel de ru ca memoria.
Pacientului i lipseste, de regul, initiativa, spontaneita-
tea si intuitia.
Agresivitatea i furia par a fi modulate de ctre
formatiunea hipocampic. Stimularea electric (la pisi-
c) a prtii hipocampice aflate cel mai aproape de amig-
dal (spre polul temporal) faciliteaz comportamentul
de tipul atacului prdtorului, iar stimularea prtii
aproapiate de aria septal inhib acest comportament.
Este important faptul c aria septal primeste aferente
de la hipocamp si trimite eferente ctre hipotalamus.
Din acest motiv, se presupune c joac rolul de releu
pentru semnalele care pleac din hipocamp pentru mo-
dularea comportamentelor agresive provocate de ctre
hipotalamus (vezi si reglarea comportamentului afectiv
Fig. 3.29. Schema bloc a conexiunilor sistemului
limbic. geile ngroate indic circuitul lui Papez
(conexiunile interne), iar sgeile subiri indic
conexiunile descrise mai recent (externe).
Curs de neuropsihologie 51
de ctre hipotalamus, cap. 3.4.5.). La om, au fost
evidentiate comportamente agresive legate de leziunile,
tumorile si epilepsia care afecteaz hipocampul. Cele
mai frecvente comportamente de acest fel sunt ostilita-
tea si actele explozive de violent fizic.
Unelefuncii endocrine sunt modulate de ctre
hipocamp, fapt realizat datorit conexiunilor cu hipota-
lamusul. Prin proiectiile sale asupra nucleului ventro-
medial, hipocampul poate inhiba secretia de ACTH
(hormon adrenocorticotrop). Se presupune c hipocam-
pul este sensibil la nivelurile hormonale si actioneaz ca
un mecanism de feedback asupra hipofizei (desigur,
prin conexiunile cu hipotalamusul fasciculul telence-
falic medial).
2) AMIGDALA
Amigdala (corpul amigdaloid, complexul
nuclear amigdaloid, nucleul amigdaloid) este o forma-
tiune alctuit din numerosi nuclei mici, fiind localizat
imediat inferior de cortexul cerebral al lobului tempo-
ral (fig. 3.19, 3.27). Prezint numeroase conexiuni
bidirectionale cu hipotalamusul, precum si cu alte arii
ale sistemului limbic. Functional, amigdala se mparte
n dou regiuni nucleare : 1) nucleii corticomediali (n
care se termin o diviziune principal a tractului olfac-
tiv ; are atasat un nucleu de talie mic) si 2) nucleii
bazolaterali (foarte dezvoltati la primate si om si nu are
legtur cu sensibilitatea olfactiv).
Conexiunile amigdalei. Amigdala primeste
aferente de la toate regiunile sistemului limbic, precum
si de la neocortexul lobilor temporali, parietali si occipi-
tali. Primeste informatii senzoriale olfactive de la bul-
bul olfactiv si informatii senzoriale auditive si vizuale
de la ariile corticale specifice. Datorit acestor cone-
xiuni multiple, amigdala a fost numit fereastra prin
care sistemul limbic receptioneaz informatii despre
mediul nconjurtor.
Cele mai importante eferente ale amigdalei se
proiecteaz pe hipotalamus prin cile striei terminale
(care porneste din nucleii corticomediali) si calea
amidgdalofugal ventral (care porneste din nucleii
bazolaterali). De asemenea, trimite napoi informatii
ctre zonele corticale de unde primeste impulsuri vizua-
le si auditive, ctre alte zone ale sistemului limbic, ctre
talamus si ctre substanta cenusie periapeductal din
mezencefal.
Funciile amigdalei. Dintre toate componente-
le sistemului limbic, amigdala exercit cel mai puternic
control asupra proceselor viscerale (vegetative) ale
hipotalamusului. Studiile au artat c amigdala este
implicat n agresivitate si furie, hrnire, functii cardio-
vasculare, endocrine, miscri somatice si memoria
afectiv.
Stimularea electric a amigdalei producereactii
asemntoare cu cele produse de stimularea electric a
hipotalamusului : cresterea sau scderea tensiunii
arteriale si a pulsului, a secretiei gastrointestinale,
mictiune, dilatarea pupilelor, piloerectie si secretia
diferitilor hormoni ai adenohipofizei (n special FSH,
LH si ACTH).
Micrile involuntare provocate de stimularea
electric a amigdalei cuprind : ridicarea capului sau a
ntregului corp, miscri circulare, diferite tipuri de
miscri asociate cu mirosirea si nghitirea alimentelor
(linsul buzelor, masticatia, salivatia).
Reactii de agresivitate i furie asemntoare cu
cele prezentate n unitatea despre hipotalamus (3.4.5.)
pot fi provocate prin stimularea anumitor nuclei.
Reactii sexuale, cum ar fi erectia, miscrile
copulatorii, ejacularea, ovulatia, contractiile uterine sau
travaliul prematur, pot fi, de asemenea, provocate.
La oameni, stimularea electric poate evoca
diferite tipuri de amintiri cu ncrctur afectiv. Deci,
amigdala este implicat nmemoria afectiv.
Patologia amigdalei. Distrugerea experimen-
tal (la maimute) a portiunilor anterioare a lobilor
temporali (care includ amigdalele) induce modificri
comportamentale reunite sub numele de sindromul
Klver-Bucy
1
. La animal, se manifest prin : lipsa
reactiei de team la orice, curiozitate extrem fat de
orice, uitare rapid, tendinte orale (bag tot ce gseste
n gur si chiar ncearc s nghit), hipersexualitate
(ncearc s copuleze cu orice fiint, indiferent de
vrst, sex sau specie). Leziuni similare sunt ntlnite
rar la om, iar simptomele sunt destul de asemntoare.
Epilepsia temporal (de lob temporal) pare a fi
legat si de leziunile amigdalei. ns nu poate fi atribui-
t n totalitate acesteia sau unei alte componente a
lobului temporal. Criza epileptic se datoreaz unui
dezechilibru ntre excitatie si inhibitie la nivelul unor
zone nucleare, care provoac o descrcare excesiv a
neuronilor. n epilepsia temporal se constat urmtoa-
rele simptome :
1) modificri viscerale : disconfort epigastric (n partea
de sus a abdomenului) si toracic, great, palpitatii,
paloare sau nrosire facial, tahipnee (accelerare a
ritmului respiratiei) sau apnee (ntreruperea
temporar a respiratiei), salivatie excesiv, eructatie
(rgial), defecatie si mictiune involuntar ;
2) modificri afective : team (de la foarte usoar la
foarte intens), sentiment de singurtate, depresie,
reactii de aprare si, mai rar, emotii pozitive (vese-
lie, voiosie, excitatie erotic) ;
3) iluzii perceptive (de tip distorsiune) : vizuale,
auditive, olfactive (mai rar) ;
4) tulburri mnezice (de memorie) : senzatie de nstr-
inare (nerecunoasterea obiectelor, evenimentelor),
senzatie de familiaritate (deja-vu).
1
Nepredat.
Curs de neuropsihologie 52
3.5.2. Nucleii bazali
Nucleii bazali (ganglioni bazali, ganglia), ca si
cerebelul, fac parte din sistemul motor extrapiramidal,
care mpreun cu sistemul piramidal este responsabil de
controlul motor. De fiecare parte a creierului, ganglionii
bazali sunt reprezentati de nucleul caudat, putamen,
globus pallidus (fig. 3.19, 3.30), substanta neagr (fig.
3.12) si nucleul subtalamic. Nucleul caudat si putame-
nul formeaz neostriatumul, globus pallidus (paleostri-
atum) si putamenul formeaz nucleul lentiform, iar
neostriatumul si nucleul lentiform (deci cei trei nuclei
caudat, putamen si palid) formeaz corpul striat. Corpul
striat include nucleii bazali dispusi n interiorul
emisferelor cerebrale, ceilalti doi nuclei fiind localizati
n subtalamus (nucleul subtalamic) si n mezencefal
(substanta neagr). Majoritatea fibrelor senzoriale si
motorii care fac legtura ntre cortexul cerebral si
mduva spinrii trec prin spatiul dintre nucleii striati,
numit capsula intern (fig. 3.19).
Nucleul caudat (fig. 3.30) are forma literei C si
este dispus de-a lungul ventriculului lateral. n partea
rostral a talamusului se gsestecapul nucleului caudat,
partea cea mai voluminoas a nucleului caudat. Poste-
rior, capul se ngusteaz si se continu cu corpul
nucleului caudat. Ultima component, coada nucleului
caudat, este cea mai subtire, se arcuiste n partea
posterioar a talamusului si ajunge la amigdal.
Nucleul lentiform (putamen +globus pallidus,
fig. 3.30) are forma unei piramide cu baza triunghiular,
aflat n dreptul insulei, de care este desprtit de
capsula extern si claustrum (o lam de substant cenu-
sie cu functie necunoscut). Vrful piramidei se afl n
apropiata vecintate a talamusului.
Pentru mai multe detalii despreglobus pallidus
si desprenucleul subtalamic vezi cap.3.4.4., iar pentru
mai mult detalii despresubstanta neagr vezi cap.3.2.3.
Conexiunile nucleilor bazali.
1
Putem vorbi de
trei tipuri de conexiuni ale nucleilor bazali (fig. 3.31) :
1. Aferente provenite, n special, de la cortexul cere-
bral (virtual, toate ariile corticale proiecteaz ctre
nucleii bazali) prin capsula intern si extern si de
la talamus (de la nucleii intralaminari) ;
2. Conexiuni interne reciproce ntre striat si substanta
neagr, ntre globus pallidum si nucleul subtalamic
(tot reciproce) si o proiectie masiv neostriato-
pallidal ;
3. Eferente : neostriatumul trimite, n special ctre
talamus si scoarta cerebral, iar paleostriatumul c-
tre talamus, nucleul rosu, formatiunea reticulat si
oliva bulbar ; dup cum se poate vedea n fig.
3.31, principalele eferente pornesc din paleostria-
tum, celelalte facnd sinaps la nivelul acestuia.
1
Nepredate.
Fig. 3.30. Localizarea nucleilor striai (nucleii bazali din
interiorul emisferelor cerebrale).
Fig. 3.31. Schema bloc conexiunilor nucleilor bazali.
Nucleii bazali sunt notai cu litere ngroate. Se poate
observa i funcia motorie a acestor nuclei, datorat
acestor conexiuni.
Curs de neuropsihologie 53
Funciile nucleilor bazali.
1
Dup cum amin-
team la nceputul acestei unitti, nucleii bazali fac parte
din sistemul extrapiramidal, un sistem secundar al con-
trolului miscrilor (sistemul principal este cel pirami-
dal, alctuit din tracturile corticospinale). De regul,
nucleii bazali au fost asociati cu functia demodulare a
initierii micrilor (realizat de ctre cortexul motor),
ns acestia sunt implicati si n alte circuite cortico-
subcorticale, care par a modula aspecte non-motorii
ale comportamentului. Aceste circuite paralele pornesc
din diferite zone ale cortexului, stimuleaz componente
specifice din nucleii bazali si talamus si se termin na-
poi n scoarta cerebral. Circuitele non-motorii ale
nucleilor bazali sunt : circuitul prefrontal (cu rol n
plnuirea constient a miscrilor), circuitul limbic (cu
un rol presupus n modularea emotiilor) si circuitul
oculomotor (cu rol n modularea miscrilor oculare).
Cile celor patru circuite sunt prezentate n fig. 3.32.
Similaritatea anatomic dintre cele trei circuite
non-motorii si cel motor sugereaz c si functionarea
lor este asemntoare. De exemplu, circuitul prefrontal
poate modula initierea si ncetarea unor functii cogniti-
ve, cum ar fi plnuirea, atentia si memoria de lucru (de
scurt durat). Analog, circuitul limbic poate modula
strile afective si motivatia. De asemenea, tulburarea
functiilor cognitve si afective prezente n boala
Parkinson si Huntington (vezi patologia) poate fi
datorat disfunctiilor acestor circuite non-motorii.
Deoarece circuitele de feedback care au traiect
1
La curs se preciza doar c functiile sunt legate de realizarea
unor actiuni instinctive si n stabilitatea fondului postural.
de la cortex spre nucleii bazali si napoi la cortex con-
tinneuroni GABA-ergici, acestea sunt circuite defeed-
back negativ, avnd efect inhibitor, deoarece GABA
(acidul gamaaminobutiric) este principalul neurotrans-
mittor inhibitor (vezi si boala Huntington). Acest
feedback negativ ofer stabilitate sistemelor pentru
controlul motor. Un rol asemntor are i DA
(dopamina), care face legtura ntre substanta neagr,
nucleul caudat si putament (vezi si boala Parkinson).
Alti neurotransmittori eliberati la nivelul nucleilor ba-
zali sunt : ACh (acetilcolina), care face legtura dintre
cortex si nostriatum, norepinefrin, 5-HT (serotonin),
enkefalin, glutamat etc.
Patologia nucleilor bazali. n linii mari,
lezarea circuitului motor poate duce la disfunctii de tip
hiperkinetic, (diskinezie miscri involuntare de tip
coree, atetoz, hemibalism, tremor, ticuri si hipotonie
muscular mai mult sau mai putin accentuat) sau
hipokinetic (bradikinezie lentoarea miscrilor si
hipertonie muscular rigiditate muscular).
Mirile de tip coreiform (coreea) sunt miscri
involuntare ample, care apar brusc si se insereaz prin-
tre miscrile voluntare. Ele se nsotesc, de regul, de o
hipotonie muscular exagerat (membre balante).
Coreea poate fi de tip Sydenham (acut) sau de tip
Huntington (cronic). Coreea Sydenham apare n copi-
lrie sau adolescent (7-13 ani), este asociat cu tul-
burri cardiace, evolueaz ctiva ani, dup care se vin-
Fig. 3.32. Schema bloc a circuitelor n care sunt implicai nucleili bazali.
Curs de neuropsihologie 54
dec). Despre coreea Huntington vom vorbi mai mult.
Atetoza const n miscri lente, de tip vermi-
form (ca viermele), continue, stereotipice, care afectea-
z muschii extremittilor si, uneori, muschii fetei si g-
tului. Sunt accentuate de emotii si miscri voluntare si
dispar n timpul somnului.
(Hemi)Balismul este o miscare involuntar vio-
lent, de tip balistic care cuprinde membrele contrala-
terale leziunii. De regul, cuprinde si muschii proxi-
mali ai membrelor, iar tulburrile de tonus lipsesc.
Tremorul, cea mai comun form de diskinezie,
este caracterizat de miscri involuntare ritmice, oscila-
torii n raport cu un punct fix. Afecteaz, de regul,
extremittile si, mai rar, capul si gtul. Tremorul poate
fi fiziologic (7-11 Hz) sau patologic. Tremorul patolo-
gic poate fi : tremor de repaus (3,5-7 Hz), tremor
postural (6-11 Hz) si tremor intentional saukinetic (3-
7 Hz). Tremorul fiziologic este slab sesizabil cu ochiul
liber, dar se poate amplifica datorit emotiilor, obose-
lii, substantelor psihoactive, hipoglicemiei etc.
Ticurile sunt miscri bruste, rapide, (de regul)
stereotipe, ce implic simultan mai multe grupe muscu-
lare. De obicei, apar n jurul ochilor si gurii si se pot
extinde ctre gt si umeri. Ticuri pot fi de tipul mic-
rilor simple (clipit, umflarea nrilor, ridicarea si cobo-
rrea umerilor etc.), micrilor complexe (dat din cap,
srit), sunete simple (ghrm, ltrat) si sunete complexe
(sughit, mrit, ecolalie repetarea cuvintelor). Ticuri-
le sunt specifice copilriei si, de regul, nu persist mai
departe. Dac totusi persist, se diminueaz n intensi-
tate si frecvent.
Coreea este provocat de lezri mici si multiple
ale putamenului, atetoza de leziunile pallidumului, iar
balismul de leziunile subtalamusului.
Tulburrile complexe datorate leziunilor nuclei-
lor bazali sunt boala Parkinson, boala Huntington si
sindromul Tourette. S le analizm pe rnd.
Boala Parkinson (paralizia agitant) este a doua
cea mai ntlnit boal degenerativ a SN dup
Alzheimer si cea mai important afectiune a nucleilor
bazali. Descris n anul 1817 de ctre J ames Parkinson
(1755-1824) sub numele deparalizie agitant, aceast
boal este caracterizat de o serie desimptome motorii,
uneori fiind nsotit si dedement. Simptomele motorii
includ tremor de repaus (3-6 Hz), dificultti mari la
initierea micrilor (bradikinezie, care poate merge,
uneori, pn la akinezie lipsa aproape total a
miscrilor) si hipertonie (rigiditate) muscular, n spe-
cial n zonele extremittilor si gtului. n medie, boala
Parkinson apare ntre 50 si 70 de ani si estefatal n
10- 20 de ani.
Tremorul involuntar (de repaus), specific n
Parkinsonism, e prezent mereu n starea de veghe, spre
deosebire de tremorul intentional provocat de lezarea
cerebelului (vezi si patologia cerebelului cap. 3.3.).
Este amplificat de emotii, oboseal, stress si anxietate
si diminuat de miscrile voluntare. Afecteaz mai ales
extremittile si arat de parc pacientul ar numra bani.
Bradikinezia este adesea mai neplcut dect
celelalte dou simptome motorii, deoarece, n formele
severe (akinezie), bolnavul trebuie s si concentreze
toat energia pentru a efectua chiar si o miscare simpl,
cum ar fi a ridica mna.
Hipertonia muscular este caracterizat de
rezistent la miscrile psive, care afecteaz att mus-
chii agonisti, ct si pe cei antagonisti. Fenomenul de
roat dintat este caracterizat de modificri periodice
ale tonusului muscular datorate tremorului adiacent si
poate fi observat si simtit la miscarea extremittilor.
Pe lng aceste simptome, se mai pot observa :
imposibilitatea de a pstra postura erect pentru mai
mult timp, aplecarea capului nainte, nghetri
temporare, aparitia rapid a somnolentei, tulburri de
vorbire si de scris (usoare), tulburri vegetative (hiper-
salivatie, constipatie) s.a.
Tulburrile cognitive (dementa) implic afecta-
rea memoriei de lucru (scurt durat), capacittii de a
plnui, nlturrii rspunsurilor nonpertinente, sponta-
neittii etc. Tulburrile afective includ aplatizarea afec-
tiv (indiferenta), depresie asociat cu lipsa de activita-
te, stereotipia activittii (intentionat), apatie etc.
Tulburrile motorii se datoreaz pierderii
progresive de neuroni dopamingergici (care produc
DA) din substanta neagr. Nucleul caudat si putame-
nul, inhibati, n stare normal de ctre DA din substanta
neagr, devin hiperactivi, transmitnd ncontinuu
impulsuri excitatorii ctre aproape toti muschii corpu-
lui. De asemenea, mai ales pentru bradikinezie, se crede
c responsabil este si distrugerea neuronilor dopami-
nergici dinsistemul limbic. Cauza distrugerii neuronilor
dopaminergici este necunoscut. Exist anumite trata-
mente care amelioreaz simptomele bolii Parkinson
(medicamentele L-Dopa si L-Deprenyl, transplant de
neuroni dopamingergici functioneaz cteva luni etc).
Boala Huntington (coreea Huntington sau coreea
cronic) a fost descris pentru prima dat n 1872, de
ctre George Huntington (1850-1916), pe baza unor
pacienti ai bunicului si tatlui su. De obicei, era con-
fundat cu tulburri psihice, cum ar fi schizofrenia sau
tulburarea sociopat, datorit simptomelor psihice
extrem de importante. Coreea Huntington este o afec-
tiune ereditar care debuteaz la 30-40 de ani. Este
Curs de neuropsihologie 55
caracterizat de deficiente motorii, cognitive si compor-
tamentale. Este o boal degenerativ cecauzeaz moar-
tea n 10-20 de ani.
Coreea (componenta motorie a bolii) se carac-
terizeaz prin miscri bruste, izolate la nceput, care
ulterior cuprind tot corpul. Cauza fiziologic a acestor
micri este dat de distrugerea majorittii neuronilor
GABA-ergici (care secret acid gamaaminobutiric) din
nucleul caudat i putamen si ai neuronilor colinergici
(care secret acetilcolin) din numeroase alte arii cere-
brale. n mod normal, terminatiile axonale ale neuroni-
lor GABA-ergici inhib globus pallidus si substanta
neagr si, desigur, lipsa acestei inhibitii determin
activarea excesiv a muschilor de ctre acesti nuclei.
Dementa din boala Huntington pare a nu fi cau-
zat de distrugerea neuronilor GABA-ergici, ci coliner-
gici, n special a celor din cortexul cerebral (care este
implicat n functiile cognitive).
Uneori, poate fi prezent si atetoza.
Defectul genetic care provoac Huntington este
localizat pe bratul scurt al cromozomului 4. Nu se
cunoaste tratament.
Sindromul Tourette a fost descris pentru prima
dat n anul 1884, de ctre George Albert douard
Brutus Gilles de la Tourette sau, pe scurt, Gilles de la
Tourette (1857-1904). Este un sidrom rar si sever, care
implic multiple ticuri : strnut, sforit, coprolalie
(limbaj obscen), ecopraxie (imitatia unor acte), ecolalie
(tendinta de a repeta cuvinte sau propozitii auzite
recent). Ticurile sunt adesea nsotite de tulburri com-
portamentale, cum ar fi tulburarea obsesiv-compulsiv,
lipsa controlului impulsurilor si lipsa atentiei. Primul
simptom, un tic motor, apare ntre doi si 14 ani.
Baza neurofiziologic a sindromului Tourette
este dat de afectarea neuronilor dopaminergici ai
nucleului striat.
Sindromul Tourette este o afectiune neuropsihi-
atric tratat cu Haloperidol, un agent care blocheaz
receptorii de DA.
3.5.3. Scoara cerebral
Scoarta cerebral (cortexul cerebral) reprezint
etajul cel mai superior al SNC, acoperind emisferele
cerebrale si fiind brzdat de girusuri si scizuri (fig.
3.24-3.26). Este o structur aprut trziu, din punct de
vedere filogenetic, iar cortexul cerebral uman reprezint
apogeul dezvoltrii filogenetice. Desi reprezint, de
departe, cea mai important component a SNC, func-
tiile sale sunt cel mai putin cunoscute. Se cunosc, ns,
efectele provocate de lezarea scoartei sau de stimularea
specific a anumitor zone, astfel stim c aici are loc
integrarea si procesarea cognitiv (perceptia, gndirea
etc.). Numrul neuronilor scoartei cerebrale nu se cu-
noaste precis, literatura fiind plin de estimri ce merg
de la 2,6 pn la 100 de miliarde ! Celulele gliale sunt
cam de vreo zece ori mai multe. Aria cortical reprezin-
t aproximativ 220 cm
2
, iar volumul este de aproxima-
tiv 300 cm
3
.
Cele mai multeaferente ale cortexului cerebral
provin de la talamus. Altele, n numr mai mic, provin
de la diverse zone subcorticale (trunchiul cerebral,
diencefal etc.) sau corticale, care pot avea origine n
aceeasi emisfer (fibrele de asociatie) sau n emisfera
opus (corpul calos).
Neuronii cortexului cerebral pot avea legturi
cu alti aproximativ 10.000 de neuroni. Sunt mprtiti n
dou mari categorii :
1) Neuroni piramidali : mici (cu efecte tonice), mijlo-
cii sau giganti (Betz), acestia din urm ajungnd
pn acolo nct pot fi observati cu ochiul liber ;
reprezint circa 66% din populatia neuronal
cortical ; mpreun cu neuronii fuziformi, dau
nastere aproape tuturor fibrelor eferente ale
cortexului ;
2) Neuroni stelati sau granulari au de obicei axoni
scurti, si functioneaz, n principal, ca interneuroni
care transmit impulsuri nervoase numai pe distante
scurte n interiorul cortexului cerebral ; unii sunt
excitatori, elibernd glutamat, iar altii sunt inhibi-
tori, elibernd GABA (cap. 2.4.) ; pot fi n form
de cos, fuziformi, orizontali, nevrogliformi si
granulari (celulele lui Martinotti) ; 33 % din total.
n functie de structura celular, cortexul cere-
bral se mparte n:
- paleocortex sau allocortex : contine dou straturi
de celule stratul extern (granular), format din
neuroni mici senzitivi si stratul intern (piramidal),
format din neuroni motori mijlocii si mari ;
- neocortex sau izocortex : contine sase straturi
delimitate ntre ele n functie de mrimea, forma si
numrul de neuroni, de densitatea fibrelor
mielinizate si de functiile specifice (pentru detalii
vezi fig. 3.33 si tabelul 3.3.).
Functiile, superioare si complexe, ale cortexului
cerebral se datoreaz organizrii celulare a acestuia,
care difer de la o regiune la alta. Aranjamentul celular
al oricrei regiuni corticale poate fi subdivizat n unitti
functionale numite coloane corticale (fig. 3.34).
numrul de neuroni este estimat pentru fiecare coloan
Curs de neuropsihologie 56
n jurul a 2500, dintre care aproximativ 100 sunt
piramidali (vezi si ultimul paragraf de la pag. 7).. Acesti
neuroni piramidali reprezint eferentele unei coloane,
iar aferentele provin de la alte coloane (fibre de
asociatie) sau de la arii senzoriale periferice.
Scoarta cerebral este mprtit ntr-o multitu-
dine de arii functionale (Broadmann a gsit cca. 50),
cum ar fi aria motorie, aria somatoestezic, aria vizual
etc. ns despre acestea vom vorbi cndva, n viitor...
Tabelul 3.3. Straturile neocortexului, celulele i terminaiile nervoase pe care le conin i funciile specifice, acestea din
urm fiind notate cu litere nclinate.
I Stratul molecular (plexiform, zonal)
Celule nervoase mici dispuse orizontal i multe celule gliale (n special
astrocite). Strat de asociaie, n principal.
II Stratul granular extern
Multe celule stelate i puine celule piramidale. Axonii se duc n jos spre
III i IV aici se integreaz mesajele senzoriale transmise prin fibrele care
pornesc de la nucleii talamici periferici .
III Stratul piramidal extern
Neuroni piramidali (mijlocii i mari) cu o dendrit lung care ajunge la
stratul molecular, iar axonii o iau n jos, strbat celelalte straturi i
trund n substana alb subcortical (neuroni de proiecie). O parte din
ei revin n aceeai emisfer, iar ceilali trec n emisfera opus (neuroni de
asociaie).
IV Stratul granular intern
ngust, conine neuroni stelai i civa piramidali al doilea sediu cortical
al sensibilitii (aferene de la nucleii talamici nespecifici).
V Stratul piramidal intern
Conine cei mai muli neuroni piramidali gigani (Betz), dar i mai mici.
Dendritele lor urc pn la I, iar axonii se duc n jos, la centrii motori
subcorticali i medulari (trimit comenzi pentru miri voluntare) stratul
mirii voluntare.
VI Stratul multiform (fuziform)
Neuroni cu forme i prelungiri variate(fuziforme, triunghiulare, ovoide,
Martinotti etc.), cu legturi subcorticale. La nivelul acestui strat se
realizeaz integrarea senzorial, formndu-se percepia.
Fig. 3.33. Aspectul laminar al neocortexului (cele ase
straturi de celule). Prin tehnica colorrii Golgi se pot
observa celulele i terminaiile lor, prin tehnica Nissl
doar corpii celulari, iar prin tehnica Weigert doar
terminaiile nervoase.
Fig. 3.34. Schema simplificat a unei coloane corticale.
Curs de neuropsiholgie 57
4. METODE I TEHNICI DE INVESTIGARE N
NEUROPSIHOLOGIE
Pentru a cerceta relatia dintre psihic si creier,
exist trei mari categorii de metode : metode
fiziologice, metode anatomice si metode psihologice.
2.1. Metode i tehnici fiziologice
Avnd dou modalitti de transmitere a informatiei
(electric si chimic), SN permite utilizarea a dou
seturi de tehnici de investigare : stimularea si
nregistrarea electric si stimularea si nregistrarea
chimic.
Stimularea electric este o tehnic ce
urmreste mimarea activittii naturale a creierului si
este cunoscut de foarte mult timp (a fost aplicat, la
om, de ctre Bartholow, n 1874, cercettorul obtinnd
date asemntoare cu cele de la stimularea pe animale).
Stimularea localizat se realizeaz cu ajutorul
electrozilor cu un singur fir, care pot fi att de subtiri
nct s permit stimularea unui singur neuron. Socurile
electroconvulsive reprezint stimularea electric cea
mai intens si este folosit uneori n tratamentul
bolnavilor depresivi. Tehnica electrozilor implantati a
furnizat date valoroase cu privire la fiziologia
creierului.
Pentru studii comportamentale se utilizeaz
electrozi mai mari, care activeaz sute sau mii de celule
nervoase. S-a artat c gradul de complexitate si
specificitate al rspunsurilor provocate este direct
proportional cu nivelul ierarhic la care se situez zona
stimulat. Stimularea de la distant (telemetric) a
relevat date aberante. Comportamentul natural uman nu
poate fi nlocuit de stimulare, acesta necesitnd
stimularea coordonat a prea multor puncte la diverse
nivele cerebrale. Utilizarea acestui procedeu ca
instrument de control sau de manipulare
comportamental este conditionat de rezolvarea
prealabil a unor probleme serioase de ordin etic, politic
si social.
nregistrarea electric este o metod ce
permite nregistrarea activittii electrice naturale a
creierului si este realizat, de asemenea, cu ajutorul
electrozilor, prin diverse tehnici electrofiziologice
(electroencefalografia, electrocorticografia, potentialele
evocate, stereoelectroencefalografia, electroence-
falografia cuantificat). nregistrarea poatea fi fcut pe
un singur neuron sau pe populatii de mii de neuroni
(cum este n cazul potentialelor evocate).
Electroencefalografia (EEG) este cea mai
frecvent utilizat n practica clinic curent si se
realizeaz cu ajutorul unor electrozi aplicati pe scalp,
dup diferite montaje, pentru a acoperi toat suprafata
capului. Totusi, prin
acest mijloc nu
acoperim dect prtile
laterale ale emisferelor
cerebrale, regiunea
median si cea
inferioar a cortexului
nefiind cuprinse. EEG
este extrem de usor si
rapid de realizat, nu
produce senzatii nepl-
cute si nu are contra-
indicatii.
La om, prima nregistrare a ritmurilor
bioelectrice a fost realizat de ctre psihiatrul german
Hans Berger (1924), ntr-o lucrare publicat n 1929.
Semnalul EEG este, regul, de origine cortical,
provenind din sumatia potentialelor postsinaptice ale
neuronilor, n special a celor piramidali. Traseul ritmic
al acestuia este dependent de integritatea functional a
structurilor subcorticale. Aceast activitate continu a
milioane de neuroni apare n dou variante :
sincronizat si desincronizat.
Cu ajutorul EEG se obtin informatii despre
patru tipuri de ritmuri (biocurenti) cerebrale : u (alfa),
(beta), A (delta), 0 (teta) (fig. 4.2.).
Fig. 4.1. Persoan la EEG.
Curs de neuropsihologie 58
Ritmul o are o frecvent de 8-13 Hz, o durat de
80-120 ms si o amplitudine de 25-100 V. Se
nregistreaz n zonele posterioare ale creierului
adult n stare de veghe, n stare de repaos cu ochii
nchisi si se atenueaz sau se blocheaz la
deschiderea ochilor (este denumit ritmul relaxrii).
Orice excitatie (luminoas, psihosenzorial, efort
de atentie, activitate psihic) anihileaz ritmul u
(reactie de oprire).
Ritmul este constituit din unde cu frecventa de
14-25 Hz si o amplitudine de 10-30 V (n medie
jumate din u). Foarte neregulate si dispersate
printre ritmurile u, ele sunt mascate si pot fi puse
n evident numai dup suprimarea ritmurilor u
printr-un excitant psihosenzorial. Aceste unde apar
n zonele temporale si frontale. Este un ritm de
activare, este mai putin labil si dispare la excitatiile
tactile.
Ritmul 0 este constituit din frecvente de 4-7 Hz si
amplitudine de 30-70 V. Apare mai ales la copii
(n jurul a patru ani, este ritmul dominant), dar
poate fi prezent si la adulti sau, n caz de
somnolent, la nivelul regiunilor temporale. Este
generat de ctre formatiunile subcorticale. Valorile
mari ale ritmului 0 indic procese patologice n
straturile cerebrale profunde. Normal, nu trebuie s
depseasc 10-15% din totalul undelor nregistrate.
Ritmul A este constituit din unde cu frecvent de
0,5-3 Hz si amplitudine de 50-100 V. Apare n
special la copiii mici (<4 ani), dar si la adulti n
timpul somnului (Non-REM). Poate fi monomorf
(asemntor u, poate indica perturbri ale
activittii electrice subcorticale) sau polimorf
(asemntor , poate indica tulburri ale activittii
electrice corticale).
Marea utilitate a EEG este n studiul si diagnosticul
epilepsiei, care se manifest prin descrcri electrice
anormale, hipersincrone, excesive si intermitente, la
nivelul cortexului cerebral.
Electrocorticografia se refer la nregistrarea
activittii electrice cerebrale, cu ajutorul electrozilor
aplicati direct pe cortexul cerebral, dup efectuarea
unor craniotomii.
Potentialele evocate sunt reactii electrice evocate n
SN de stimuli externi specifici. n clinic, se utilizeaz
frecvent potentiale evocate vizuale, auditive si
somatosenzitive. n vederea acestui scop, se prezint
subiectului stimulul de studiat, n mod repetitiv, dup
care se face o medie a rspunsurilor, care permite
degajarea potentialelor de voltaj mic, marcate pe EEG.
Deoarece creierul are o activitate permanent, este
imposibil a deosebi un potential evocat singular de
activitatea de fond. Rspunsurile apar la cteva
milisescunde de la aplicarea stimulului, latenta
depinznd de localizarea exact a electrodului
nregistrator. Cel mai adesea, potentialele evocate sunt
preluate din cortex cu ajutorul electrozilor plasati direct
pe scalp, dar ele pot fi nregistrate si cu ajutorul unei
singure unitti.
Stereoelectroencefalografia cu electrozi
implantati n profunzime a permis efectuarea unor studii
asupra miscrilor anormale n cursul talamotomiilor.
nregistrrile si stimulrile talamusului ne-au adus
informatii neuropsihologice cu privire la rolul
structurilor acestuia n limbaj si memorie. nregistrrile
la nivelul prtii ventromediane a lobilor frontali, la
nivelul prtii centrale a lobilor temporali si a regiunii
din jurul ventriculului III au contribuit la explorarea
substratului anatomic al emotiilor umane. De asemenea,
au permis si stabilirea unor legturi ntre anumite
fenomene comportamentale si localizarea lor anatomic
(de exemplu, agresivitate si hipocamp). Astzi, metoda
este utilizat n studiul epilepsiei si n tratamentul bolii
Parkinson.
Electroencefalografia cuantificat se foloseste
pentru studiile lateralizare ale proceselor psihice, la
studiile asupra asimetriei functionale (deseori
contestate), iar n psihofarmacologie, pentru depistarea
si clasificarea noilor psihotrope. Este putin folosit,
momentan.
Stimularea i nregistrarea chimic se face cu
ajutorul substantelor halucinogene (mescalin, opium,
morfin, LSD etc.) si a medicamentelor ce au ca efect
modificarea comportamentului si tririlor psihologice
(antidepresive, antiaxioase, analgezice etc.). Pe msura
acumulrii experientei privind neurotrasmiterea
sinaptic la nivel cerebral, a devenit posibil
controlarea efectelor comportamentale ale
Fig. 4.2. Ritmurile EEG.
Curs de neuropsihologie 59
medicamentelor n conjunctie cu transmittorii si
receptorii sinaptici, dezvoltndu-se astfel
psihofarmacologia sau asa-numita farmacologie
comportamental.
Psihofarmacologia cuprinde dou mari domenii.
Primul se ocup de mecanismele de actiune ale
medicamentelor utilizate n clinica psihiatric, n timp
ce al doilea are n vedere utilizarea medicamentelor ca
unelte de investigare a functiilor cerebrale legate de
comportament.
n principiu, orice substant medicamentoas ce
actioneaz asupra creierului, actioneaz si asupra
comportamentului este o metod de stimulare chimic.
Actiunile acestora difer : n timp ce unele pot actiona
la nivelul sinapsei, altele pot actiona asupra membranei
neuronale prin scdere excitabilittii acesteia
(anestezicele), iar altele interfereaz cu metabolismul
neurotransmittorilor.
Analiza lichidului cefalorahidian (cerebro-
spinal) se face prin punctii, realizate n zona lombar
sau suboccipital. Cnd se face analiza lichidului
cefalorahidian, urmrim: compozitia sa chimic si
prezenta unor particule virale sau microbiene. Este
principala modalitate de confirmare sau infirmare a
diagnosticului de meningit, meningoencefalit sau
hemoragie cerebral si nu numai.
Metode fiziologice globale. Meritul trecerii de
la cercetarea artificial-segmetar la cea sistemic-
natural, a activittii reflexe, i revine lui Pavlov.
Introducnd notiunile de reflex conditionat, semna-
lizare si legtur temporar, el a pus bazele unui capitol
nou, fundamental al fiziologiei SN, care a adus o
contributie decisiv la ntelegerea obiectiv-stiintific a
raportului psihic-creier. Metoda conditionrii a dus la
determinarea legilor generale ale formrii si
consolidrii reflexelor conditionate de diferite tipuri
(simple, de diferentiere, de ntrziere, stereotipii
dinamice etc.). Tot cu aceast metod, s-au putut
identifica si determina propriettile naturale ale celor
dou procese fundamentale (excitatia si inhibitia),
respectiv forta, echilibrul si mobilitatea. Dup cum
stim, acestea au dus la elaborarea teoriei
temperamentale dinamice, dar si la analiza
neurodinamic a altor structuri de personalitate.
Skinner perfectioneaz metoda conditionrii,
prin introducerea termenului de conditionare
instrumental sau operant. Acest fapt relev c, n
conditiile mediului natural de viat, att animalul, ct si
omul se confrunt frecvent cu situatii problematice de
diferite grade de complexitate, care se interpun ntre
motivatia subiectului si obiectul corespunztor
satisfacerii ei. Pentru aceasta, omul sau animalul trebuie
s-si formeze pe loc acte comportamentale adecvate
surmontrii obstacolelor ce se impun ntre organism si
obiect. Aceste acte se numesc instrumentale sau
operante, deoarece ele mijlocesc si faciliteaz atingerea
obiectivului adaptativ final. Analiza lor furnizeaz
informatii cu privire la modul general de functionare a
creierului si la organizarea comportamentului.
2.2. Metode i tehnici anatomice
Metoda leziunilor anatomice. Leziunea
nseamn nlturarea unei mici portiuni dintr-o structur
anatomic, cu ajutorul bisturiului, aspiratiei, injectrii
cu alcool, refrigerare focal, prin implantarea granulelor
radioactive, coagulare, electric, tehnica neurotoxinelor
etc. La om, leziunea mai poate fi produs si de o boal
neurologic sau de o interventie chirurgical. Ablatia
(care nu prea se mai foloseste) se refer la ndeprtarea
unei ntregi componente anatomice (cortexul, trunchiul
cerebral etc.). Lobotomia (rar utilizat) se refer la
distrugerea fizic a unui lob cerebral, iar leucotomia are
drept scop sectionarea unor ci comune dintre diferite
portiuni ale creierului.
Metoda leziunilor anatomice vizeaz relatia
dintre o regiune circumscris a creierului lezat si
deficientele cognitiv-comportamentale ce apar n urma
lezrii. Mai concret, aceasta vizeaz determinarea
specializrii functionale a diferitelor structuri si zone
ale creierului si stabilirea legturii dintre acestea si
diferitele functii psihice.
Metode neuropatologice. Orice afectiune
patologic a creierului provoac tulburri
corespunztoare n plan psihocomportamental. De
aceea, metodele neuropatologice se folosesc de un
material servit ca pe tav, cercettorul avnd direct o
leziune provocat si trebuind s stabileasc doar
corelatul psihocomportamental (cum au fcut
Wernicke, Broca, J ackson, Penfield s.a.). Totusi, nu
trebuie s ne pripim n a face corelatii, deoarece
localizarea tulburrilor unei functii psihice oarecare nu
este totuna cu localizarea functiei respective normale.
Trebuie consemnate cel putin trei momente ale
dinamicii tulburrii : tabloul pre-operator, tabloul
postoperator imediat si tabloul postoperator ndeprtat.
La baza acestei metode trebuie s stea anumite
principii dup care s fie dirijat : principiul adecvrii,
principiul gradatiei, principiul obiectivittii, principiul
functionalittii dinamice.
Curs de neuropsihologie 60
Principiul adecvrii. Pentru explorarea, n
scopuri de diagnostic, a fiecrui proces psihic trebuie
folosite sarcini specifice, diversificate si diferentiate,
care s permit diferentierea tipurilor de tulburri.
Principiul gradatiei. Presupune c explorarea
oricrui proces psihic trebuie s urmeze, pe de-o parte
drumul de la simplu la complexe si, pe de alt parte, s
tin seama de diferentele interindividuale (vrst, sex,
pregtire etc.).
Principiul obiectivittii. Presupune : a) stabili-
rea, pentru fiecare prob, a unor indicatori si parametri
de ordin cantitativ si calitativ exteriorizabili si msura-
bili ; b) posibilitatea de verificare a datelor n situatii
similiare ; c) predictabilitatea verificabilitatea prin
alte metode clinice (fiziologice, electrofiziologice, ima-
gistice, biochimice etc.).
Principiul functionalittii. Orice prob neuro-
psihologic aplicat cu metoda neuropatologic trebuie
s aib o constructie problematic (trebuie s pun
subiectul n contextul unei mici procesualitti).
Examinarea neuroradiologic se foloseste n
general pentru depistarea sindroamelor, dar, mai nou,
metodele neuroradiologice (se mai gsesc si sub numele
de imagistic cortical), mai concret, imagistica prin
rezonant magnetic (IRM sau MRI, n englez), se
foloseste si pentru a determina ariile cerebrale activate
n timpul solicitrii unor procese psihice. Neuroradio-
logia dispune de radiografie, angiografie, tomografie
computerizat (TC) si IRM.
Radiografia permite examinarea obiectiv a
oaselor si a unor tesuturi moi din corpul uman. Radio-
grafia foloseste razele X, descoperite din greseal de
ctre Wilhelm Conrad Rntgen, n 1895, pentru care a
primit primul Premiu Nobel pentru Fizic, n 1901. Pe
cliseele simple (imagini standard) ale craniului pot fi
evidentiate leziuni osoase, corpi strini intracranieni si
calcifieri intracraniene patologice determinate de
leziuni, precum craniofaringiomul, scleroza tuberoas,
meningiomul, oligodendrogliomul, cisticercoza, boala
Sturge-Weber s.a. Radiografia nu trebuie repetat des,
deoarece razele X sunt nocive i pot determina aparitia
meningioamelor.
Angiografia cerebral (Egas Moniz, 1927)
implic radiografii craniene, efectuate n secvent
rapid, de fractiuni de secund, n momentul injectrii
n artere a unei substante de contrast. Se foloseste, nc,
pentru evidentierea tumorilor, pentru evidentierea
deplasrilor, pentru ghidarea interventiilor exploratorii
ale neurochirurgului etc. Angiografiei clinice i se
adaug, acum, angiografia prin rezonant magnetic.
Tomografia computerizat (tomodensitometria
sau scanografia) a revolutionat neurologia, neurochirur-
gia si neuroradiologia, permitnd vizualizarea direct a
creierului. Este o metod sofisticat, care combin
tehnologia calculatorelor cu razele Rntgen. TC se
bazeaz pe iradierea creierului din toate unghiurile cu
raze X si pe relevarea diferentelor de densitate, att
normale, ct si patologice. Un computer reconstruieste
sectiunile bidimensionale oferind, n final, o vedere
tridimensional, att a structurilor cerebrale normale,
ct si a celor patologice. Este des utilizat n cazurile de
traumatism cranian, n leziunile craniene infectioase, n
leziunile degenerative cerebrale n depistarea precoce a
tumorile maligne sau benigne, primitive sau
metastazice, iar n neuropsihiatrie, se foloseste pentru
sprijinirea ipotezei diagnostice de dement non-
organic.
Imagistica prin rezonant magnetic (IRM) este
cea mai sofisticat unealt imagistic din toate
timpurile. Aceasta nu produce efecte secundare nocive,
pentru c nu utilizeaz raze X si nici substante radioac-
tive, de contrast. IRM utilizeaz variatiile semnalelor
produse de proton n corp n conditiile n care capul
pacientului este introdus ntr-un camp magnetic
puternic (fig. 4.3.). Tehnica permite msurarea efecte-
lor magnetice foarte slabe asupra tesuturilor, ceea ce
face posibil vizualizarea anatomiei organelor situate n
profunzime, opace, dar, mai ales, a activittii circuitelor
cerebrale. Se pot realiza sectiuni virtuale care permit
vizualizarea detaliilor structurii masei cerebrale (mate-
ria cenusie, materia alb) cu precizie milimetric.
Imagistica functional prin rezonant
magnetic (fIRM saufMRI englez) const n utiliza-
rea IRM pentru a vizualiza regiunile anatomice cerebra-
le unde se concentreaz activittile cognitive sau
motorii. Tehnica fIRM permite stabilirea unei hrti a
functionrii creierului uman, cu o rezolutie anatomic si
temporal neegalat pn acum (fig. 4.3.). Mecanismul
de semnalizare a
activittii cerebrale se
bazeaz pe cresterea
cantittii de snge n
zona respectiv si de
cresterea oxigenrii ace-
lui snge, consecint a
nevoilor metabolice
crescute n zon. n
momentul de fat sunt
efectuate mai ales studii
privind stimulrile vizuale, ale memoriei, ale limbajul-
ui, ale auzului si n cutarea unui focar epileptogen. Mai
Fig. 4.3. Imagini fIRM.
Curs de neuropsihologie 61
nou, tehnica fIRM este folosit si n detectarea
minciunilor.
Tomografia cu pozitroni (PET positron
emission tomography) foloseste un dispozitiv prin care
se autoradiografiaz tesuturile, prin injectarea unor
substante de contrast (radioizotopi) : oxigen 15,
nitrogen 13, carbon 11, glucoz radioactiv (oxigenul si
glucoza sunt consumate cu predilectie de ctre creier).
Cu ajutorul unui computer se obtine o imagine de o
precizie remarcabil (fig.
4.4.). Din punctul de vedere
al cercetrii, PET se foloses-
te n studiul zonelor cerebra-
le implicate n vorbire, dar
si pentru studierea altor
functii cognitive sau pentru
msurarea debitului sangvin
cerebral.
Fig. 4.4. Scanarea creierului
cu PET.
2.3. Metode psihologice
Investigarea neuropsihologic nu trebuie s se rezume
la metodele neurologice de investigare (analizate mai
sus), ci trebuie s le implice, desigur, si pe cele
psihologice. Metodele psihologice obiective sunt
centrate pe relevarea, nregistrarea si analiza ct mai
veridic si mai fin a structurii si dinamicii actelor
comportamentale. Acest lucru nu este deloc simplu, rea-
lizarea celei mai simple miscrii sau reactii putnd
implica actiunea unui numr mare de componente
psihice (de exemplu, cnd ne aducem aminte un cuvnt,
se zice c am folosit mecanismul de reactualizare al
memoriei, ns acesta implic aportul gndirii, precum
si asocieri verbale, n cazul cuvintelor cu sens, dar si
asocieri imagistice, n aproape orice fel de memorare).
Astfel, complexitatea extraordinar a sistemului psihic
uman a fcut necesar elaborarea unei game largi de
metode si procedee de cercetare. Vom analiza patru
dintre aceste metode, care par a fi esentiale pentru
investigarea neuropsihologic : metoda analizei
produselor activittii, metoda testelor, metoda experi-
mentului si de laborator, metoda genetic si comparat.
Metoda analizei produselor activitii se
foloseste pentru cunoasterea motivatiilor, intereselor si
disponibilittilor creatoare ale persoanei. n acest scop,
se recurge la sarcini de desen, compunere, constructie
de obiecte etc.
Metoda testelor a fost introdus n neuropsi-
hologie pentru a determina nivelul dezvoltrii mintale
generale, cu scopul de a distinge anormalitatea de
normalitate si realizarea mai eficient a orientrii si
selectiei profesionale. Testul poate fi utilizat att ca
instrument de diagnosticare, ct si ca instrument de
cercetare neuropsihologic. O problem n legtur cu
testele este numrul lor imens (se estimeaz peste
10.000), ceea ce duce la dificultti de alegere si stp-
nire. Dintre cele mai cunoscute teste, amintim testul de
inteligent general Binet-Simon, testul aperceptiv
tematic (TAT), Rorsarch, 16-PF (Cattell), testul
Eysenck.
Metoda experimental i de laborator
presupune un control riguros al variabilei independente
si al conditiilor de desfsurare a experimentului. Nu mai
struim asupra acestei metode, fiind desfsurat pe larg
n cursul de psihologie experimental. Pentru detalii,
vezi M. Anitei (2007), Psihologie Experimental,
Editura Polirom.
Metoda genetic i comparat are la baz prin-
cipiul fundamental al devenirii si evolutiei, potrivit
cruia organizarea psihic nu trebuie interpretat ca
ceva dat, predeterminat si imuabil, ci ca un produs al
evolutiei filogenetice si ontogenetice. Pentru a ntelege
un proces psihic aflat la un anumit nivel de structurare
si integrare, trebuie s cercetm etapele genetice ante-
rioare pe care le-a parcurs. Prin aplicarea metodei
genetice au putut fi descoperite legi fundamentale ale
organizrii psihice, fr de care astzi este de
neconceput elaborarea unei teorii psihologice generali-
zate, cu adevrat stiintifice. Printre acestea, o importan-
t metodologic major pentru neuropsihologie o au
urmtoarele : legea dezvoltrii ; legea interactiunii
dialectice dintre cauzele si factorii externi, pe de-o
parte, si conditiile interne (inclusiv factorii ereditari), pe
de alt parte ; legea stadialittii ; legea heterocroniei ;
legea heteronomiei.
Legea devenirii sustine c psihicul devine, se
formeaz n timp, deci, postuleaz caracterul derivat,
devenit al psihicului, precum si faptul c devenirea nu
este ncheiat, este optimizabil si variabil. Un aspect
esential pe care ni-l relev legea dezvoltrii este sensul
ascendent al transformrilor, desfsurarea lor de la
simplu la complex, de la organizare slab la organizare
mai bun.
Legea interactiunii dialectice dintre cauzele i
factorii externi i conditiile interne postuleaz c
procesul devenirii si dezvoltrii sistemului psihic uman
este declansat si determinat, pe toat durata lui, de
influentele externe si mai ales de interactinea dintre
Curs de neuropsihologie 62
aceste influente si caracteristicile sistemului endocrin,
experienta anterioar si strile actuale de necesitate
(motivatia). De-a lungul vietii, omul avanseaz de la o
interactiune biologic/fiziologic ntre persoana lui si
mediu, la una din ce n ce mai psihologic, adic com-
portamentul su va deveni va deveni din ce n ce mai
bine stpnit de structurile psihice superioare.
Legea stadialittii surprinde caracterul gradual
al formrii si dezvoltrii tuturor functiilor si proceselor
psihice. Legea a fost demonstrat de cercettori celebri
ce au investigat cu precdere anumite laturi ale vietii
psihice. Piaget a demonstrat stadialitatea inteligentei,
Wallon a demonstrat stadialitatea dezvoltrii afective,
Freud a demonstrat stadialitatea centrrii si satisfacerii
libidoului, ajungnd la o stadialitatea psihosexual,
Erikson a demonstrat asa-numita stadialitate
psihosocial (confruntarea dintre trebuintele individului
si ceea ce-i ofer societatea). Important de sublinat este
faptul c trecerea de la un stadiu la altul n planul dez-
voltrii psihocomportamentale este sustinut si,
implicit, corelat cu trecerea de la un nivel inferior la
unul superior n organizarea structural-functional a
creierului (de exemplu, mielinizarea).
Legea heterocroniei postuleaz c diferitele
functii, procese si structuri psihice se constituie si se
consolideaz la momente diferite de timp (de exemplu,
mecanismele perceptive se formeaz si se dezvolt
nainte de cele ale gndirii). Piaget a artat c stadiile
nu se omit si nu se inverseaz deoarece ntre ele exist
interactiuni necesare genetic, adic, un stadiu actual
pregteste aparitia unui nou stadiu, asigurnd conditiile
ca n stadiul urmtor s se produc o serie de
transformri.
Legea heteronomiei reflect caracterul eterogen
si contradictoriu al dezvoltrii psihice la nivel
intraindividual si interindividual. Prin urmare, ea are
dou postulate de baz : 1) n interiorul sistemului
psihic al unei persoane, diferite componente ating
niveluri diferite de dezvoltare (vz mai bun dect auz,
gndire abstract mai bun dect concret etc.) si 2) una
si aceeasi functie sau structur psihic are niveluri
diferite de dezvoltare la doi sau mai multi indivizi.
Metoda logico-matematic, logico-formal i
cibernetic se foloseste n situatiile n care ne
propunem s modelm un anume proces psihic.
Curs de neuropsihologie 63
BIBLIOGRAFIE
Albu, I, Georgia, R (2004), Anatomie clinic, ed. a III-a,
Bucuresti, Editura ALL.
Anitei, M (2007), Psihologie experimental, Iasi, Polirom.
Arseni, C (1980) (coord.), Tratat de neurologie, vol. II.
Sindroamele neurologice, Bucuresti, Editura
Medical.
Barker, RA, Barasi, S, Neal, MJ (1999), Neuroscience at a
Glance, Oxford, Blackwell Science Ltd.
Botez, M (coord.) (1996), Neuropsihologie clinic i
neurologia comportamentului, Bucuresti, Editura
Medical.
Brazis, PW, Masdeu, J C, Biller, J (2007), Localization in
Clinical Neurology, 5
th
ed., Lippincott Williams &
Wilkins.
Carlson, N (2005), Foundations of Physiological
Psychology, 6th ed., Boston, Pearson Education
Inc.
Dale, P, Augustine, GJ, Fitzpatrick, D, Hall, WC, LaMantia,
A-S, McNamara, JO, Williams, SM (eds.),
Neuroscience, 3
rd
ed., Massachusetts, Sinauer
Associates Inc.
Dnil, L, Golu, M (2006), Tratat de neuropsihologie, vol.
I+II, Bucuresti, Editura Medical.
Ellis, H (2006), Clinical Anatomy, 11
th
ed., London,
Blackwell Publishing.
Evans-Martin, FF (2005), Your Body How It Works The
Nervous System, New York, Chelsea House
Publishers.
Fix, JD, High-Yield Neuroanatomy, 2
nd
ed., Lippincott
Williams & Wilkins.
Goetz, CG (2007), Textbook of Clinical Neurology, 3
rd
ed.,
Philadelphia, Elsevier Inc.
Goldstein, LH, McNeil, J E (eds.) (2004), Clinical
Neuropsychology, The Atrium, Southern Gate,
Chichester, West Sussex, John Wiley and Sons Ltd.
Greenstein, B, Greenstein, A (2000), Color Atlas of
Neuroscience : Neuroanatomy and Neurophy-
siology, New York, Thieme.
Gross, R (2005), Psychology Science of Mind and
Behavior, 5
th
ed., Dubai, Hodder Arnold.
Guyton, AC, Hall, J E (2006), Tratat de fiziologie a omului,
ed. a 11-a, Bucuresti, Editura Medical Callisto.
Haines, DE (2007), An Atlas of Structures, Sections and
Systems, 6
th
ed., Lippincott Williams & Wilkins.
Haines, DE (ed.) (2007), Fundamental Neuroscience for
Basic and Clinical Applications, 3
rd
ed.,
Philadelphia, Elsevier Inc.
Kahle, W, Frotscher, M (2003), Color Atlas of Human
Anatomy, Vol. 3 : Nervous System and Sensory
Organs, 5
th
ed., New York, Thieme.
Kolb, B, Whishaw, IQ (2003), Fundamentals of Human
Neuropsychology, 5
th
edition, New York Freeman-
Worth.
Mare, V (1976), Mecanismele neurofiziologice ale activittii
psihice, in Roca, Al. (coord.), Psihologie
general, Bucuresti, Editura Didactic si
Pedagogic.
Netter, FH, Craig, J A, Perkins, J (2002), Atlas of
Neuroanatomy and Neurophysiology, special
edition, Austin, Texas, Icon Custom
Communications.
Netter, FH (2006), Atlas of Human Anatomy, 4
th
ed.,
Philadelphia, Elsevier-Saunders.
Niculescu, CT, Voiculescu, B, Nit, C, Crmaciu, R.,
Slvstru, C, Ciornei, C (2007), Anatomia i
fiziologia omului : compendiu, Bucuresti, Editura
Corint.
Pocock, G, Richards, CD (2006), Human Physiology : The
Basis of Medicine, 3
rd
ed., Oxford University Press.
Rohkamm, R (2004), Color Atlas of Neurology, New York,
Thieme.
Ropper, AH, Brown, RH (2005), Adams and Victor's
Principles of Neurology, 8
th
ed., New York,
McGraw-Hill.
Scanlon, VC, Sanders, T (2007), Essentials of Anatomy and
Physiology, 5
th
ed., Philadelphia, F.A. Davis
Company.
Siegel, A, Sapru, HN (eds.) (2006), Essential Neuroscience,
Lippincott Williams & Wilkins.
Smith, EE, Nolen-Hoeksema, S, Fredrickson, BL, Loftus,
GR (2005), Introducere n psihologie, Bucuresti,
Editura Tehnic.
Smith, CUM, Elements of Molecular Neurobiology, 3
rd
ed.,
The Atrium, Southern Gate, Chichester, West
Sussex, John Wiley and Sons Ltd.
The Society for Neuroscience (2002), Brain Facts A Primer
on the Brain and Nervous System, 4
th
ed., China,
Everbest Printing Company.
Van de Graaff, K (2001), Human Anatomy, 6th ed., New
York, McGraw-Hill.
Young, PA, Young PH (2000), Neuroanatomie general i
clinic, Bucuresti, Editura Medical Callisto.
Waugh, A, Grant, A (2004), Ross and Wilson - Anatomy and
Physiology in Health and Illness, 9
th
ed., Spain,
Elsevier Inc.
www.anatomyatlases.org, accesat iulie 2008.
www.neuroanatomy.wisc.edu, accesat iulie 2008.
neurocomputing.org, accesat iulie 2008.
braininfo.rprc.washington.edu, accesat iulie 2008.
You might also like
- NeuropsihologieDocument50 pagesNeuropsihologieAnna DnlNo ratings yet
- Istoria psihologiei științifice: De la nașterea psihologiei până la neuropsihologie și cele mai actuale domenii de aplicareFrom EverandIstoria psihologiei științifice: De la nașterea psihologiei până la neuropsihologie și cele mai actuale domenii de aplicareRating: 5 out of 5 stars5/5 (2)
- Neuropsihologie Versiune P. StudentiDocument37 pagesNeuropsihologie Versiune P. StudentiMaximBelenchiiNo ratings yet
- Interventii neobisnuite: Modificări ale cadrului, metodei și relației în psihoterapie și psihanalizăFrom EverandInterventii neobisnuite: Modificări ale cadrului, metodei și relației în psihoterapie și psihanalizăNo ratings yet
- NeuropsihologieDocument125 pagesNeuropsihologieGeorgiana100% (6)
- Neuropsihologie Curs IntegralDocument80 pagesNeuropsihologie Curs Integralsasha_st100% (11)
- Disonanta cognitiva. Noi perspective asupra unei teorii fundamentale pentru psihologieFrom EverandDisonanta cognitiva. Noi perspective asupra unei teorii fundamentale pentru psihologieNo ratings yet
- PsihoneurofiziologieDocument220 pagesPsihoneurofiziologieCorina Enache100% (2)
- Psihologia Sinelui Prelegerile de la Institutul de Psihanaliza din ChicagoFrom EverandPsihologia Sinelui Prelegerile de la Institutul de Psihanaliza din ChicagoNo ratings yet
- Curs NeuropsihologieDocument104 pagesCurs Neuropsihologieapi-3729398100% (8)
- Prinde-i inainte de a cadea Noi abordari in tratamentul prabusirii psihiceFrom EverandPrinde-i inainte de a cadea Noi abordari in tratamentul prabusirii psihiceRating: 5 out of 5 stars5/5 (2)
- Neuropsihologie TemaDocument18 pagesNeuropsihologie TemaIoana Ana Maria Porutiu100% (4)
- NeurostiinteDocument92 pagesNeurostiinteNeamtiu Adrian100% (2)
- NeurostiinteDocument1 pageNeurostiintevanwyngardenNo ratings yet
- Tulburari ale Sinelui O abordare axata pe tipurile de personalitateFrom EverandTulburari ale Sinelui O abordare axata pe tipurile de personalitateNo ratings yet
- Formarea Pentru PsihoterapieDocument517 pagesFormarea Pentru PsihoterapieRuxandra Ioana Florea91% (22)
- NEUROPSIHOLOGIE: elementele de bază ale problemeiFrom EverandNEUROPSIHOLOGIE: elementele de bază ale problemeiRating: 5 out of 5 stars5/5 (1)
- CURS Neuropsihol 2Document55 pagesCURS Neuropsihol 2Andrei Băcanu100% (2)
- Bazele Genetice Ale Comportamentului UmanDocument49 pagesBazele Genetice Ale Comportamentului UmanduddywNo ratings yet
- Conștiința: Descoperirea etapelor minții: de la conștient la inconștient, de la influența ritmurilor biologice la somn și viseFrom EverandConștiința: Descoperirea etapelor minții: de la conștient la inconștient, de la influența ritmurilor biologice la somn și viseRating: 5 out of 5 stars5/5 (1)
- Larousse - PsihanalizaDocument59 pagesLarousse - Psihanalizacioreli100% (4)
- Grile NeuroDocument21 pagesGrile NeuroIrina EvaNo ratings yet
- Psihicul si corpul: Descoperirile și implicațiile psihologiei analiticeFrom EverandPsihicul si corpul: Descoperirile și implicațiile psihologiei analiticeRating: 5 out of 5 stars5/5 (1)
- Mintea ta secreta Cunoaste-ti si asuma-ti inconstientulFrom EverandMintea ta secreta Cunoaste-ti si asuma-ti inconstientulRating: 5 out of 5 stars5/5 (1)
- Orientari moderne in psihoterapie si consiliere psihologicaFrom EverandOrientari moderne in psihoterapie si consiliere psihologicaRating: 4.5 out of 5 stars4.5/5 (4)
- Psihoterapia psihodinamica Practica bazata pe dovezi empiriceFrom EverandPsihoterapia psihodinamica Practica bazata pe dovezi empiriceRating: 1 out of 5 stars1/5 (1)
- ICD-10-CM Studii de caz pentru viitorii psihologi clinicieni Probleme psihologice si comportamentaleFrom EverandICD-10-CM Studii de caz pentru viitorii psihologi clinicieni Probleme psihologice si comportamentaleRating: 5 out of 5 stars5/5 (3)
- Lumea interpersonala a copilului O perspectiva bazata pe psihanaliza si psihologia dezvoltariiFrom EverandLumea interpersonala a copilului O perspectiva bazata pe psihanaliza si psihologia dezvoltariiNo ratings yet
- Psihoterapiile cognitive si comportamentale in tulburarile de personalitate Aplicatii practice si noi directiiFrom EverandPsihoterapiile cognitive si comportamentale in tulburarile de personalitate Aplicatii practice si noi directiiRating: 5 out of 5 stars5/5 (2)
- PSICOLOGIE, DEPRESIUNE ȘI DISTURBARE UMORALĂ: înțelegerea mecanismelor de bazăFrom EverandPSICOLOGIE, DEPRESIUNE ȘI DISTURBARE UMORALĂ: înțelegerea mecanismelor de bazăRating: 5 out of 5 stars5/5 (1)
- Ce-ar face Freud in locul tau? Cum ar rezolva cei mai mari psihoterapeuti problemele tale cotidieneFrom EverandCe-ar face Freud in locul tau? Cum ar rezolva cei mai mari psihoterapeuti problemele tale cotidieneRating: 5 out of 5 stars5/5 (1)
- Sinele: Bazele si implicatiile unui concept central din psihologia analiticaFrom EverandSinele: Bazele si implicatiile unui concept central din psihologia analiticaNo ratings yet
- Psihosomatica. Teorii, cazuri, descriere medicalaFrom EverandPsihosomatica. Teorii, cazuri, descriere medicalaRating: 4.5 out of 5 stars4.5/5 (3)
- Înțeles, sens și referință în filosofia limbajului și logica filosoficăFrom EverandÎnțeles, sens și referință în filosofia limbajului și logica filosoficăNo ratings yet
- Psihopatologia vietii cotidiene Despre uitare, greseli de vorbire, superstitie si eroareFrom EverandPsihopatologia vietii cotidiene Despre uitare, greseli de vorbire, superstitie si eroareRating: 5 out of 5 stars5/5 (1)
- Limbajul uitat O introducere in intelegerea viselor, basmelor si miturilorFrom EverandLimbajul uitat O introducere in intelegerea viselor, basmelor si miturilorRating: 5 out of 5 stars5/5 (4)
- Pacienti narcisici, terapeuti incepatori: Conceptualizare, tratament si gestionarea contratransferuluiFrom EverandPacienti narcisici, terapeuti incepatori: Conceptualizare, tratament si gestionarea contratransferuluiRating: 5 out of 5 stars5/5 (2)
- Alfred Adler și psihologia individuală în noul mileniu: Strategii, principii și modele operaționale care stau la baza gândirii fondatorului psihologiei individualeFrom EverandAlfred Adler și psihologia individuală în noul mileniu: Strategii, principii și modele operaționale care stau la baza gândirii fondatorului psihologiei individualeNo ratings yet
- Atacurile de panica: Un program pentru a evita capcanele paniciiFrom EverandAtacurile de panica: Un program pentru a evita capcanele paniciiRating: 5 out of 5 stars5/5 (1)
- Distincția dintre falsificare și respingere în problema demarcației la Karl PopperFrom EverandDistincția dintre falsificare și respingere în problema demarcației la Karl PopperNo ratings yet
- Introducere in interpretarea jungiana a desenelorFrom EverandIntroducere in interpretarea jungiana a desenelorRating: 5 out of 5 stars5/5 (2)