You might also like
- Cellular and Subcellular Specification of Na, K-ATPase A and B Isoforms in The Postnatal Development of Mouse RetinaDocument12 pagesCellular and Subcellular Specification of Na, K-ATPase A and B Isoforms in The Postnatal Development of Mouse RetinaJose GarciaNo ratings yet
- Are Changes in MAPK ERK Necessary or Sufficient For Entrainment in Chick Pineal CellsDocument11 pagesAre Changes in MAPK ERK Necessary or Sufficient For Entrainment in Chick Pineal CellsJose GarciaNo ratings yet
- Margery C. Beinfeld Diana M. KorchakDocument8 pagesMargery C. Beinfeld Diana M. KorchakJose GarciaNo ratings yet
- Transcription Factors in Neuroendocrine Regulation Rhythmic Changes in pCREB and ICER Levels Frame Melatonin SynthesisDocument11 pagesTranscription Factors in Neuroendocrine Regulation Rhythmic Changes in pCREB and ICER Levels Frame Melatonin SynthesisJose GarciaNo ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (844)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5810)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- Human Eye 1Document13 pagesHuman Eye 1GUNJAN SHARMANo ratings yet
- Tuberculoma OF THE Brain (Revisited)Document7 pagesTuberculoma OF THE Brain (Revisited)Nyoman SuryaNo ratings yet
- MSE DVD Booklet Oct 2011 High ResolutionDocument21 pagesMSE DVD Booklet Oct 2011 High ResolutionNurul Hidaya HamliNo ratings yet
- Vascular Malformations of The Central Nervous SystemDocument9 pagesVascular Malformations of The Central Nervous SystemFranklin Aranda100% (2)
- Neurological Disorders ProjectDocument3 pagesNeurological Disorders ProjectDavid Vander HaakNo ratings yet
- The Senses: Eugene Flor Lindawan Ulpindo, MSNDocument67 pagesThe Senses: Eugene Flor Lindawan Ulpindo, MSNayoNo ratings yet
- Chapter 13 Nervous System in Mammals - Worksheet (Answers)Document2 pagesChapter 13 Nervous System in Mammals - Worksheet (Answers)api-3728508100% (2)
- KinurerinDocument19 pagesKinurerinKanita BakijaNo ratings yet
- Hand ExaminationDocument16 pagesHand ExaminationRezaul KarimNo ratings yet
- Head InjuryDocument50 pagesHead InjuryThiagarajah RaviNo ratings yet
- Nursing Care Management 101 Health Assessment: Procedural Checklist inDocument5 pagesNursing Care Management 101 Health Assessment: Procedural Checklist inKyle Randolf CuevasNo ratings yet
- Memory: Sta. Teresa CollegeDocument4 pagesMemory: Sta. Teresa Collegeaudree d. aldayNo ratings yet
- 9 Multiple SeclrosisDocument38 pages9 Multiple SeclrosisAhmed aljumailiNo ratings yet
- The Neurophenomenology of Gesture in The PDFDocument18 pagesThe Neurophenomenology of Gesture in The PDFabrahamzapruderNo ratings yet
- نسخة Lecture - PAIN Assessment & ManagementDocument52 pagesنسخة Lecture - PAIN Assessment & Managementjsoal100% (1)
- Meet Your Happy ChemicalsDocument9 pagesMeet Your Happy ChemicalsIvan MontNo ratings yet
- Self-Projection and The Brain: Randy L. Buckner and Daniel C. CarrollDocument9 pagesSelf-Projection and The Brain: Randy L. Buckner and Daniel C. CarrollJOHNATIELNo ratings yet
- Science PortfolioDocument81 pagesScience PortfolioVismaya ViswanathanNo ratings yet
- Current Protocols in Toxicology (2005) BM OCR 7.0-2.6 LotBDocument2,759 pagesCurrent Protocols in Toxicology (2005) BM OCR 7.0-2.6 LotBHamida MidaNo ratings yet
- Test Bank For Guyton and Hall Textbook of Medical Physiology 13th Edition by John e Hall 005 2Document6 pagesTest Bank For Guyton and Hall Textbook of Medical Physiology 13th Edition by John e Hall 005 2Matthew Anderson97% (29)
- Perceptual DisordersDocument3 pagesPerceptual DisordersAkshayaaNo ratings yet
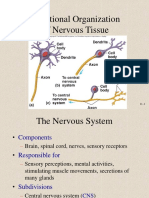
- Functional Organization of Nervous TissueDocument28 pagesFunctional Organization of Nervous TissueIssa JoyahaNo ratings yet
- Individual Project - PresentationDocument55 pagesIndividual Project - PresentationTan Kar YeowNo ratings yet
- Pe1 FlexibilityDocument7 pagesPe1 FlexibilityErielle SibayanNo ratings yet
- Sensory For The SLP: Cari Ebert, MS, CCC-SLPDocument30 pagesSensory For The SLP: Cari Ebert, MS, CCC-SLPDave RoinNo ratings yet
- ABR ShortDocument50 pagesABR ShortLau Teik BengNo ratings yet
- Rahu and Eighth HouseDocument4 pagesRahu and Eighth HouseSatyanneshi ERNo ratings yet
- Substance Misuse Disorders: Dr. Ravi PaulDocument19 pagesSubstance Misuse Disorders: Dr. Ravi PaulRavi PaulNo ratings yet
- Cogpsy Reviewer 1 2Document18 pagesCogpsy Reviewer 1 2MANALANG JULIA ALEXISNo ratings yet