You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5810)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (844)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Computer Simulation: Regulation of Gene Expression - / 30 PtsDocument3 pagesComputer Simulation: Regulation of Gene Expression - / 30 PtsBeatriz Amabelle GAYAPANo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Human Physiology Final ExamDocument7 pagesHuman Physiology Final ExamXuân ViNo ratings yet
- Scar Management in Burn Injuries Using Drug Delivery and 2018 Advanced DrugDocument20 pagesScar Management in Burn Injuries Using Drug Delivery and 2018 Advanced DrugDIOGENESNo ratings yet
- Pathophysiology of Insulin ResistanceDocument8 pagesPathophysiology of Insulin ResistanceCamilo VidalNo ratings yet
- Lab 3.5 FinaleDocument16 pagesLab 3.5 FinaleAlberto Anderson100% (4)
- Biogear Tissue Homogenizer Prep: Samples PreparationsDocument1 pageBiogear Tissue Homogenizer Prep: Samples PreparationsNormenNo ratings yet
- Inqaba Biotec Company Profile 2022 FinalDocument12 pagesInqaba Biotec Company Profile 2022 FinalOlufela Yomi-LayinkaNo ratings yet
- The Cell Nattapat2021Document119 pagesThe Cell Nattapat2021Poochita SongsriNo ratings yet
- Lecture 2 - CytogeneticsDocument19 pagesLecture 2 - CytogeneticsHusssainNo ratings yet
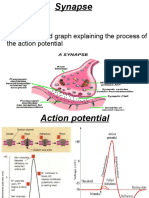
- Synapse 1214834359710231 8 PDFDocument11 pagesSynapse 1214834359710231 8 PDFMuhammad AbdullahNo ratings yet
- Restriction EnzymesDocument14 pagesRestriction EnzymesKnowinter0% (2)
- Hey GuysDocument2 pagesHey GuysJaidelNo ratings yet
- Soal VPJ Edit FinalDocument8 pagesSoal VPJ Edit FinalIntan Rita SavitriNo ratings yet
- Ferroptosis in Different Pathological ContextsDocument16 pagesFerroptosis in Different Pathological ContextsSpore FluxNo ratings yet
- Vivas in Surgical Pathology For DM 1: What Is A Neoplasm? Define Neoplasia?Document11 pagesVivas in Surgical Pathology For DM 1: What Is A Neoplasm? Define Neoplasia?Giovanni HenryNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument15 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsArya ChowdhuryNo ratings yet
- Weber K. y Osborn M. 1969. The Reliability of Molecular Weight Determinations by DodecylDocument7 pagesWeber K. y Osborn M. 1969. The Reliability of Molecular Weight Determinations by DodecylEsaú Bojorquez VelazquezNo ratings yet
- Biology Data: D N A V T N B, BDocument3 pagesBiology Data: D N A V T N B, Bapara_jitNo ratings yet
- Biology Passing Material KAYAKA PU COLLEGE KALABURAGIDocument23 pagesBiology Passing Material KAYAKA PU COLLEGE KALABURAGIRAJAGOPAL93% (15)
- Sarcopenia, Frailty and Their Prevention by ExerciseDocument22 pagesSarcopenia, Frailty and Their Prevention by ExerciseJoe PolancoNo ratings yet
- 260002-Pharmaceutical Microbiology and Biotechnology-IIDocument2 pages260002-Pharmaceutical Microbiology and Biotechnology-IIchintoo4696No ratings yet
- Robert Clive Landis, Kim R. Quimby and André R. Greenidge: Review ArticleDocument9 pagesRobert Clive Landis, Kim R. Quimby and André R. Greenidge: Review ArticleΔημητρης ΦιλιογλουNo ratings yet
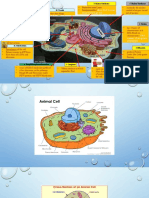
- DRAFT Animal Cell and FunctionsDocument3 pagesDRAFT Animal Cell and FunctionsSumayya KassimNo ratings yet
- II.a. Cell The Basic Unit of LifeDocument27 pagesII.a. Cell The Basic Unit of LifeDarwin NoolNo ratings yet
- Formal Report For BiochemDocument4 pagesFormal Report For BiochemJaz AlcalaNo ratings yet
- Protein Intake PTC8 PDFDocument110 pagesProtein Intake PTC8 PDFvNo ratings yet
- Cell - The Unit of Life, Cell Cycle and Cell Division - Practice Sheet 02 - Yakeen NEET 2024Document5 pagesCell - The Unit of Life, Cell Cycle and Cell Division - Practice Sheet 02 - Yakeen NEET 2024ankitfrutyNo ratings yet
- Engineering Enzyme Microenvironments For Enhanced BiocatalysisDocument10 pagesEngineering Enzyme Microenvironments For Enhanced BiocatalysisTUSHAR DASHNo ratings yet
- Mechanism of Blood Clotting Extensic Pathway Factors Affecting Blood ClottingDocument18 pagesMechanism of Blood Clotting Extensic Pathway Factors Affecting Blood ClottingRaunak TripathiNo ratings yet
- Unidad II Micro PDFDocument71 pagesUnidad II Micro PDFAlfredo RodriguezNo ratings yet