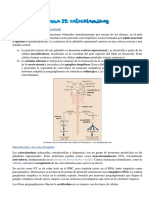
You might also like
- Metabolismo y efectos adversos de los CannabinoidesFrom EverandMetabolismo y efectos adversos de los CannabinoidesNo ratings yet
- Adrenalina y NoradrenalinaDocument8 pagesAdrenalina y Noradrenalinajuan migel herrrera fernandez100% (1)
- ADRENALINA FisiologíaDocument4 pagesADRENALINA FisiologíaXimena MedinaNo ratings yet
- 1.2.1.1. Adrenalina y NoradrenalinaDocument11 pages1.2.1.1. Adrenalina y NoradrenalinaEd ChávezNo ratings yet
- Biosíntesis de Las CatecolamínasDocument6 pagesBiosíntesis de Las CatecolamínassiscupNo ratings yet
- Neurotransmisión AdrenérgicaDocument16 pagesNeurotransmisión AdrenérgicaDiane BarrigaNo ratings yet
- NoradrenalinaDocument8 pagesNoradrenalinaJesús Guzmán100% (1)
- Tarea 6 Equipo3Document12 pagesTarea 6 Equipo3Alan DominguezNo ratings yet
- Colg-Bioqui. R1 T8. 08-04-23Document3 pagesColg-Bioqui. R1 T8. 08-04-23Stewe Yefry BravoNo ratings yet
- ADRENALINADocument11 pagesADRENALINAJohan Sebastian Cardona EscobarNo ratings yet
- Drogas VasoactivasDocument80 pagesDrogas Vasoactivascarlos gordilloNo ratings yet
- 3 Adrenalina, Noradrenalina. 3ra ClaseDocument88 pages3 Adrenalina, Noradrenalina. 3ra ClaseRosa PaezNo ratings yet
- DOPAMINADocument48 pagesDOPAMINASü PQ100% (3)
- Clase 8Document7 pagesClase 8Lisbeth Adriana Lopez AriasNo ratings yet
- Fármacos ColinomimetricosDocument9 pagesFármacos ColinomimetricosRonny RomeroNo ratings yet
- Cat Eco LaminaDocument8 pagesCat Eco LaminaIrving Oswaldo Chable VazquezNo ratings yet
- Neurotransmisores PDFDocument9 pagesNeurotransmisores PDFAladino AbbasNo ratings yet
- FARMACO 5 Farmacologia Del SNC y PerifericoDocument7 pagesFARMACO 5 Farmacologia Del SNC y PerifericoBimix NoxNo ratings yet
- Aminas Simpaticomimeticas AnticonvulsivantesDocument14 pagesAminas Simpaticomimeticas AnticonvulsivantesMont PzkNo ratings yet
- Apuntes Farmacología II Parte 1 (Corregido)Document27 pagesApuntes Farmacología II Parte 1 (Corregido)Jack Torres CastilloNo ratings yet
- Farmacologia Del SNC 2020 Parte 2 Con AudioDocument29 pagesFarmacologia Del SNC 2020 Parte 2 Con AudioLucas PercenNo ratings yet
- Catecolaminas FarmacologiaDocument2 pagesCatecolaminas FarmacologiaMARIA DE LOS ANGELES CLAVO DUCLOSNo ratings yet
- Neurotransmisión AdrenérgicaDocument4 pagesNeurotransmisión AdrenérgicaSol ArayaNo ratings yet
- SEMINARIO TALLER - AntiparkinsonianosDocument8 pagesSEMINARIO TALLER - AntiparkinsonianosElva Roxana Chavez LiñanNo ratings yet
- Tema 28 - CatecolaminasDocument12 pagesTema 28 - CatecolaminasNatalia BecerraNo ratings yet
- Catecolaminas - BioquimicaDocument4 pagesCatecolaminas - BioquimicaAlvaro GarciaNo ratings yet
- NNDocument32 pagesNNLupita GutierrezNo ratings yet
- Farmacos Moduladores de La NoradrenalinaDocument5 pagesFarmacos Moduladores de La NoradrenalinaJazmin Shany StellaNo ratings yet
- Síntesis de CatecolaminasDocument3 pagesSíntesis de CatecolaminasSergio Morales100% (1)
- Aspectos Celulares y Moleculares en PsiquiatríaDocument15 pagesAspectos Celulares y Moleculares en PsiquiatríaRielc zemartinNo ratings yet
- DopaminaDocument8 pagesDopaminamenamoisNo ratings yet
- Farmacologia ExpoDocument39 pagesFarmacologia ExpoOmar Ruelas GilNo ratings yet
- NeurotransmisiónDocument11 pagesNeurotransmisiónValentina MercadoNo ratings yet
- CLASE 8 Agonistas AdrenergicosDocument41 pagesCLASE 8 Agonistas AdrenergicosGianfranco TalaveraNo ratings yet
- DopaminaDocument11 pagesDopaminaYeimy Flores CantutaNo ratings yet
- Adrenalina: Receptores Alfa AdrenérgicosDocument12 pagesAdrenalina: Receptores Alfa AdrenérgicosAbrahamTrinidadNúñezNo ratings yet
- Farm. General - Ao N⺠3Document13 pagesFarm. General - Ao N⺠3Wilder CanelonNo ratings yet
- Neurotransmisores Uam Est 2017-2Document32 pagesNeurotransmisores Uam Est 2017-2oriel pinedaNo ratings yet
- Do PaminaDocument9 pagesDo PaminaAmber ElvirNo ratings yet
- Orientadora 3Document12 pagesOrientadora 3Adri LópezNo ratings yet
- Semana 5Document13 pagesSemana 5Tatiana MarquesNo ratings yet
- Las CatecolaminasDocument17 pagesLas CatecolaminasPurpleGold175No ratings yet
- Catecolaminas 120815191953 Phpapp01Document24 pagesCatecolaminas 120815191953 Phpapp01elieliabregoNo ratings yet
- Adrenergicos YaDocument24 pagesAdrenergicos YaVania RengifoNo ratings yet
- Agonistas Adrenergicos ClaseDocument51 pagesAgonistas Adrenergicos ClaseCleysser Antonio Custodio Polar0% (1)
- FenetilaminaDocument18 pagesFenetilaminaMoises Valdivia BaldomeroNo ratings yet
- DopanimaDocument8 pagesDopanimaHenry AcevedoNo ratings yet
- CatecolaminasDocument3 pagesCatecolaminasPaula Tarapues100% (1)
- Morfologia SinapticaDocument4 pagesMorfologia SinapticaIvan Figueroa MartinezNo ratings yet
- NeurotransmisoresDocument2 pagesNeurotransmisoresJoseth Carolina SantanaNo ratings yet
- Sistema Simpático PDFDocument45 pagesSistema Simpático PDFMiko IbáñezNo ratings yet
- Do PaminaDocument14 pagesDo PaminamemowebberNo ratings yet
- UWIENER CLASE TEORIA 6 SEM 3 2024-0 Adrenergicos y Bloqueadores AdrenergicosDocument78 pagesUWIENER CLASE TEORIA 6 SEM 3 2024-0 Adrenergicos y Bloqueadores AdrenergicosRuth HuaringaNo ratings yet
- Adrenalina y NoradrenalinaDocument3 pagesAdrenalina y NoradrenalinaAlberto PumarejoNo ratings yet
- Sesión 11 Farmacologia IDocument12 pagesSesión 11 Farmacologia IayarNo ratings yet
- Do PaminaDocument7 pagesDo PaminaClaudio LazoNo ratings yet
- Los-Neurotransmisores 1Document2 pagesLos-Neurotransmisores 1davidNo ratings yet
- Catecolaminas RDocument19 pagesCatecolaminas RSandy HernandezNo ratings yet
- ADRENÉRGICODocument13 pagesADRENÉRGICODaniela NevoNo ratings yet
- Alimento y MedicamentoDocument8 pagesAlimento y MedicamentoEdwinNo ratings yet
- Drogas Adrenergicas o Simpaticomimeticas 10Document14 pagesDrogas Adrenergicas o Simpaticomimeticas 10R Martín Apresa M100% (1)
- Cap11 Drogas Simpaticolíticas o Adrenolíticas Malgor ValseciaDocument9 pagesCap11 Drogas Simpaticolíticas o Adrenolíticas Malgor ValseciaLuis Diego Caballero Espejo100% (1)
- Cap8 AnticolDocument7 pagesCap8 AnticolEdwin100% (5)
- Drogas Parasimpaticomimeticas o Colinergicas 08Document8 pagesDrogas Parasimpaticomimeticas o Colinergicas 08R Martín Apresa MNo ratings yet
- Cap5 Farmacometría Malgor ValseciaDocument8 pagesCap5 Farmacometría Malgor ValseciaLuis Diego Caballero EspejoNo ratings yet
- Cap6 SnaDocument19 pagesCap6 SnaEdwin100% (3)
- Cap1 Introducción A La Farmacología Malgor ValseciaDocument12 pagesCap1 Introducción A La Farmacología Malgor ValseciaLuis Diego Caballero EspejoNo ratings yet
- Cap3 Farmacodinamia Malgor ValseciaDocument7 pagesCap3 Farmacodinamia Malgor ValseciaLuis Diego Caballero EspejoNo ratings yet
- Matías Caiceo Fuentes - Tarea - 4Document7 pagesMatías Caiceo Fuentes - Tarea - 4Alondra Badilla MolinaNo ratings yet
- John DeweyDocument3 pagesJohn DeweyPablo AdroverNo ratings yet
- Guía GENEROS PERIODISTICOSDocument4 pagesGuía GENEROS PERIODISTICOSAnita MariaNo ratings yet
- Costos Logisticos de Aprovisionamiento PDFDocument8 pagesCostos Logisticos de Aprovisionamiento PDFluzNo ratings yet
- Prejardin OctubreDocument3 pagesPrejardin OctubreOlga lucia rodriguezNo ratings yet
- OBRA POETICA COMPLETA Por LUIS HERNANDEZ PDFDocument544 pagesOBRA POETICA COMPLETA Por LUIS HERNANDEZ PDFSiddharta2711No ratings yet
- Recuperación de La DemocraciaDocument3 pagesRecuperación de La DemocraciajavieraNo ratings yet
- ParametrizacionDocument23 pagesParametrizacionJose Luis Guillen VelascoNo ratings yet
- Archivos Venezolanos de Puericultura y Pediatría: Arch Venez Puer Ped Vol.76 No.2 Caracas Jun. 2013Document7 pagesArchivos Venezolanos de Puericultura y Pediatría: Arch Venez Puer Ped Vol.76 No.2 Caracas Jun. 2013Jackson CoronelNo ratings yet
- 7 Recursividad e InducciónDocument19 pages7 Recursividad e InducciónFlor BogadoNo ratings yet
- Sociologia Juridica PDFDocument10 pagesSociologia Juridica PDFJose Huaman Campos100% (1)
- 1° Comunicación, Quechua ChankaDocument240 pages1° Comunicación, Quechua Chankamitchell100% (1)
- Libro 1Document5 pagesLibro 1RICHAR MORENONo ratings yet
- Cartilla Rover0Document12 pagesCartilla Rover0Caami VillalbaNo ratings yet
- Normas DE LA BANDA DE MÚSICADocument3 pagesNormas DE LA BANDA DE MÚSICAErika Melissa Sena CallirgosNo ratings yet
- Parranda La Negrita Grupo 5Document2 pagesParranda La Negrita Grupo 5esio ocaña igarzaNo ratings yet
- Derecho Penal I PDFDocument11 pagesDerecho Penal I PDFdavidNo ratings yet
- No. 4 - 2003 - Rostros y Rastros - 0 PDFDocument30 pagesNo. 4 - 2003 - Rostros y Rastros - 0 PDFCamilo ErnestoNo ratings yet
- Guia3-Arturo El DragónDocument31 pagesGuia3-Arturo El DragónCarolinaNo ratings yet
- 2 2 Funcionalismo y CulturaDocument12 pages2 2 Funcionalismo y CulturanadiaNo ratings yet
- Como Crear Mapas Mentales - Tony BuzanDocument64 pagesComo Crear Mapas Mentales - Tony BuzanLuis Alberto Mendoza Muñoz100% (9)
- Proyecto RobóticaDocument14 pagesProyecto RobóticaSebastian ChAvezNo ratings yet
- El Deporte en Las Diferentes Teorías SociológicasDocument3 pagesEl Deporte en Las Diferentes Teorías SociológicasJavier FelizNo ratings yet
- Trabajo en Word - Mortalidad MaternaDocument8 pagesTrabajo en Word - Mortalidad Maternaalexis liendoNo ratings yet
- El Triangle Rosa-2Document7 pagesEl Triangle Rosa-2Ainhoa Navarro82% (17)
- Enfoque Disciplina Formativa Convive en La EscuelaDocument4 pagesEnfoque Disciplina Formativa Convive en La Escuelabontes67100% (1)
- EbionismoDocument3 pagesEbionismobernabe antonioNo ratings yet
- Destrezas MDocument82 pagesDestrezas MCAMILO ANDRES VARGAS PABONNo ratings yet
- Iniciacion Ganchillo Es PDFDocument0 pagesIniciacion Ganchillo Es PDFJackie RiveraNo ratings yet
- Trabajo Práctico 8 Salidas de InfostatDocument2 pagesTrabajo Práctico 8 Salidas de InfostatasdNo ratings yet