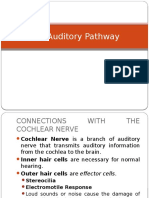
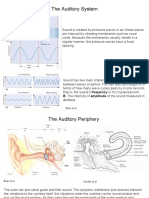
You might also like
- Mechanisms of Perception: Audition and MoreDocument24 pagesMechanisms of Perception: Audition and Morejeyn100% (1)
- The Child-Voice in Singing: Treated from a physiological and a practical standpoint and especially adapted to schools and boy choirsFrom EverandThe Child-Voice in Singing: Treated from a physiological and a practical standpoint and especially adapted to schools and boy choirsNo ratings yet
- Physiology of HearingDocument3 pagesPhysiology of HearingKC RajagopalNo ratings yet
- Sound Localization EssayDocument11 pagesSound Localization EssayscholifyNo ratings yet
- Ch. 9Document51 pagesCh. 9Sara MitteerNo ratings yet
- 0956 CH13Document12 pages0956 CH13Felipe MatheusNo ratings yet
- Do You Hear What I Hear?:: A Lecture On The Auditory SenseDocument39 pagesDo You Hear What I Hear?:: A Lecture On The Auditory Sensecherub23No ratings yet
- The Physiology of HearingDocument25 pagesThe Physiology of HearingAyi Abdul BasithNo ratings yet
- Chapter 2: Articulatory, Auditory and Acoustic Phonetics. PhonologyDocument4 pagesChapter 2: Articulatory, Auditory and Acoustic Phonetics. PhonologycalluraNo ratings yet
- Representation of Multiple Sound Locations in Primate BrainDocument19 pagesRepresentation of Multiple Sound Locations in Primate Brainacademics2014No ratings yet
- Nihms 2989Document15 pagesNihms 2989Akbar Eka PutraNo ratings yet
- Auditory and Vestibular PathwaysDocument20 pagesAuditory and Vestibular Pathwaysmoda4No ratings yet
- AD Unit-4Document38 pagesAD Unit-4Vasanth VasanthNo ratings yet
- DR Karel JindrakDocument35 pagesDR Karel Jindrakpost2vishalNo ratings yet
- Sensory and Motor SystemDocument12 pagesSensory and Motor SystemrobertoNo ratings yet
- Script 2Document6 pagesScript 2pjoyie0708No ratings yet
- Understanding the Auditory Cortex and its Role in Sound ProcessingDocument29 pagesUnderstanding the Auditory Cortex and its Role in Sound ProcessingRose RodziNo ratings yet
- Stanley norman graven auditory development and acoustic sensitivityDocument2 pagesStanley norman graven auditory development and acoustic sensitivityyoxoxec954No ratings yet
- Faal 4 - Telinga (DR - Sony)Document46 pagesFaal 4 - Telinga (DR - Sony)sarahwahyuni36No ratings yet
- Auditory ReceptorsDocument15 pagesAuditory ReceptorsJeanette RiosNo ratings yet
- Cellular Mechanisms Underlying Brain WavesDocument7 pagesCellular Mechanisms Underlying Brain WavesNick HaleyNo ratings yet
- Binaural Beats: Auditory Brainwave EntrainmentDocument6 pagesBinaural Beats: Auditory Brainwave EntrainmentBalázs EcsenyiNo ratings yet
- Musical Illusions - D.deutschDocument29 pagesMusical Illusions - D.deutschJulieta SchererNo ratings yet
- Unit 5 Bio4Er NotesDocument72 pagesUnit 5 Bio4Er NoteskgpunithaNo ratings yet
- Sensory PathwaysDocument4 pagesSensory PathwaysDaphne CallisonNo ratings yet
- Cleaning of Brain Cells by SoundDocument27 pagesCleaning of Brain Cells by SoundRadoslav Shterev100% (1)
- Binaural BeatsDocument7 pagesBinaural BeatsShubham MishraNo ratings yet
- 5.3 Hearing - Introduction To Psychology - 1st Canadian EditionDocument7 pages5.3 Hearing - Introduction To Psychology - 1st Canadian EditionWesley Z.EspinosaNo ratings yet
- Pitch Perceotion and Auditory Stream (Medical) PDFDocument16 pagesPitch Perceotion and Auditory Stream (Medical) PDF董明No ratings yet
- Loudspeaker and Listener Positions For Optimal Low-Frequency Spatial Reproduction in Listening RoomsDocument21 pagesLoudspeaker and Listener Positions For Optimal Low-Frequency Spatial Reproduction in Listening RoomsDurairajan AasaithambaiNo ratings yet
- Auditory FeaturesDocument3 pagesAuditory FeaturesShahad Qahtan AbdulameerNo ratings yet
- How Stimulation Becomes SensationDocument6 pagesHow Stimulation Becomes SensationAndrea PietersNo ratings yet
- Zatorre - Music, The Food of NeuroscienceDocument4 pagesZatorre - Music, The Food of NeuroscienceCecilia Barreto100% (2)
- ShowDocument123 pagesShowStarLink1No ratings yet
- The Auditory PathwayDocument15 pagesThe Auditory PathwayCherub SastrugiNo ratings yet
- The Oae StoryDocument24 pagesThe Oae StoryCaio LeônidasNo ratings yet
- Blindfold Improves Hearing Ability ExperimentDocument9 pagesBlindfold Improves Hearing Ability ExperimentFrancis Raymundo TabernillaNo ratings yet
- BearCh11 ACRQedtDocument7 pagesBearCh11 ACRQedtmohammadmazen281100No ratings yet
- Binaural Science PDFDocument5 pagesBinaural Science PDFRajko VojnovićNo ratings yet
- Other Sensory SystemsDocument95 pagesOther Sensory SystemsXi En LookNo ratings yet
- Anatomy and Physiology of NeuronsDocument4 pagesAnatomy and Physiology of NeuronsVan Jasper Pimentel BautistaNo ratings yet
- Module 20 Reading AssignmentDocument2 pagesModule 20 Reading AssignmentDana M.No ratings yet
- The Alchemy of Music and The BrainDocument5 pagesThe Alchemy of Music and The BrainSAW PI100% (1)
- The Auditory System: B. The Intensity of Amplitude of The Sound Measured inDocument10 pagesThe Auditory System: B. The Intensity of Amplitude of The Sound Measured inFikri RamaNo ratings yet
- AQA Biology A-level: Organisms respond to changesDocument11 pagesAQA Biology A-level: Organisms respond to changessamNo ratings yet
- Acoustic Sensors and ActuatorsDocument47 pagesAcoustic Sensors and ActuatorsGoldis RaulNo ratings yet
- 21 Howard1997Document13 pages21 Howard1997jesús buendia puyo100% (1)
- M1CHAP3Document28 pagesM1CHAP3Eilla Marie GomosNo ratings yet
- Seeing Sounds and Hearing Colors: An Event-Related Potential Study of Auditory-Visual SynesthesiaDocument14 pagesSeeing Sounds and Hearing Colors: An Event-Related Potential Study of Auditory-Visual SynesthesiaVeronika ShiliakNo ratings yet
- Pitch Perception: Arizona State University, Tempe, ArizonaDocument15 pagesPitch Perception: Arizona State University, Tempe, ArizonaFernando Flora FilhoNo ratings yet
- Yost09 PDFDocument15 pagesYost09 PDFАлесь Шлык100% (1)
- Language and The BrainDocument8 pagesLanguage and The BrainAntonio GoqueNo ratings yet
- The Vestibular System: Sensing Head MovementsDocument14 pagesThe Vestibular System: Sensing Head MovementsEslam ElsayedNo ratings yet
- 33-3 The SensesDocument19 pages33-3 The Sensesnancie8No ratings yet
- Lab Act Special SensesDocument7 pagesLab Act Special SensesJhia TorreonNo ratings yet
- Acoustic Brainwave Entrainment With Binaural BeatsDocument70 pagesAcoustic Brainwave Entrainment With Binaural BeatscesardlNo ratings yet
- SGD 15 Oktober 2019Document27 pagesSGD 15 Oktober 2019Wilson HalimNo ratings yet
- 8th NerveDocument91 pages8th NerveKinjal SharmaNo ratings yet
- Mood Disorders Short EssayDocument4 pagesMood Disorders Short EssayscholifyNo ratings yet
- 1295200672hebb's Memory Short2 EssayDocument5 pages1295200672hebb's Memory Short2 EssayscholifyNo ratings yet
- Glutamate Receptors Short EssayDocument4 pagesGlutamate Receptors Short EssayscholifyNo ratings yet
- Ectopic Pregnancy IntroDocument2 pagesEctopic Pregnancy IntroscholifyNo ratings yet
- Experimental Techniques Short Essay + NotesDocument6 pagesExperimental Techniques Short Essay + NotesscholifyNo ratings yet
- Addiction Short2 EssayDocument4 pagesAddiction Short2 EssayscholifyNo ratings yet
- The Role of Serotonin in Depression EssayDocument4 pagesThe Role of Serotonin in Depression EssayscholifyNo ratings yet
- What Is The Cellular Ion of The Cerebellum EssayDocument7 pagesWhat Is The Cellular Ion of The Cerebellum EssayscholifyNo ratings yet
- Animal Models Short Exam EssayDocument8 pagesAnimal Models Short Exam EssayscholifyNo ratings yet
- Schizophrenia 4 EssayDocument2 pagesSchizophrenia 4 EssayscholifyNo ratings yet
- What Is The Neuropharmacological Basis of Drug Addiction II EssayDocument5 pagesWhat Is The Neuropharmacological Basis of Drug Addiction II EssayscholifyNo ratings yet
- Write An Essay On Schizophrenia and The Drugs That Can Be Used To Treat It IIDocument4 pagesWrite An Essay On Schizophrenia and The Drugs That Can Be Used To Treat It IIscholifyNo ratings yet
- Opiates - EssayDocument7 pagesOpiates - Essayscholify100% (1)
- What Are The Contributions of Different Visual Cortical Areas To Analysing A Visual Stimulus EssayDocument6 pagesWhat Are The Contributions of Different Visual Cortical Areas To Analysing A Visual Stimulus EssayscholifyNo ratings yet
- Schizophrenia EssayDocument3 pagesSchizophrenia EssayscholifyNo ratings yet
- Sound Frequency Analysis EssayDocument3 pagesSound Frequency Analysis EssayscholifyNo ratings yet
- Neuronal Migration Essay For Fs - With Corrections - EssayDocument3 pagesNeuronal Migration Essay For Fs - With Corrections - EssayscholifyNo ratings yet
- 1295190621parkinson's Disease and Basal Ganglia EssayDocument4 pages1295190621parkinson's Disease and Basal Ganglia EssayscholifyNo ratings yet
- 1295188734oculomotor Control 2 - EssayDocument5 pages1295188734oculomotor Control 2 - EssayscholifyNo ratings yet
- 1295189588parkinson's Disease For Justin Boyes 2 - EssayDocument3 pages1295189588parkinson's Disease For Justin Boyes 2 - EssayscholifyNo ratings yet
- Describe The Neural Mechanisms Underlying The Stretch Reflex. How Might This Contribute To The Control of Posture and Movement?Document3 pagesDescribe The Neural Mechanisms Underlying The Stretch Reflex. How Might This Contribute To The Control of Posture and Movement?scholifyNo ratings yet
- Pain Essay - What Are The Pathways Involved in The Sensation of Pain and How Can We Manipulate Them For Our BenefitDocument5 pagesPain Essay - What Are The Pathways Involved in The Sensation of Pain and How Can We Manipulate Them For Our BenefitscholifyNo ratings yet
- 1295190511neuro Essay 4 - How Is The Information in Sound Waves Coded in The Form of Nerve Impulses by The Ear EssayDocument3 pages1295190511neuro Essay 4 - How Is The Information in Sound Waves Coded in The Form of Nerve Impulses by The Ear EssayscholifyNo ratings yet
- 1295190513neuro Essay 8 - What Is The Role of The Cerebral Cortex in Voluntary Movement EssayDocument4 pages1295190513neuro Essay 8 - What Is The Role of The Cerebral Cortex in Voluntary Movement EssayscholifyNo ratings yet
- 1295188732neuro Essay 7 - How Have Studies of Neuropharmacology Helped Our Understanding of Movement Disorders EssayDocument4 pages1295188732neuro Essay 7 - How Have Studies of Neuropharmacology Helped Our Understanding of Movement Disorders EssayscholifyNo ratings yet
- 1295188730neuro Essay 2 - Compare The Properties of Rods and Cones, and Their Retinal Pathways EssayDocument4 pages1295188730neuro Essay 2 - Compare The Properties of Rods and Cones, and Their Retinal Pathways EssayscholifyNo ratings yet
- 1295188732neuro Essay 6 - What Is The Cellular Ion of The Cerebellum How Might This Ion Relate To Its Function EssayDocument4 pages1295188732neuro Essay 6 - What Is The Cellular Ion of The Cerebellum How Might This Ion Relate To Its Function EssayscholifyNo ratings yet
- 1295188730neuro Essay 3 - Describe Some of the Diverse Types of Selectivity Shown by Visual Receptive Fields in the Retina and in the Visual Cortex and Offer Some Explanation for the Existence of This Diversity EssayDocument4 pages1295188730neuro Essay 3 - Describe Some of the Diverse Types of Selectivity Shown by Visual Receptive Fields in the Retina and in the Visual Cortex and Offer Some Explanation for the Existence of This Diversity EssayscholifyNo ratings yet
- Long Term Ion EssayDocument3 pagesLong Term Ion EssayscholifyNo ratings yet
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (811)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (3)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (4)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (515)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Wayfinding: The Science and Mystery of How Humans Navigate the WorldFrom EverandWayfinding: The Science and Mystery of How Humans Navigate the WorldRating: 4.5 out of 5 stars4.5/5 (18)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- A Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouFrom EverandA Series of Fortunate Events: Chance and the Making of the Planet, Life, and YouRating: 4.5 out of 5 stars4.5/5 (62)
- Crypt: Life, Death and Disease in the Middle Ages and BeyondFrom EverandCrypt: Life, Death and Disease in the Middle Ages and BeyondRating: 4 out of 5 stars4/5 (3)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (378)
- This Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyFrom EverandThis Is Your Brain On Parasites: How Tiny Creatures Manipulate Our Behavior and Shape SocietyRating: 3.5 out of 5 stars3.5/5 (31)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (215)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (15)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- The Invention of Tomorrow: A Natural History of ForesightFrom EverandThe Invention of Tomorrow: A Natural History of ForesightRating: 4.5 out of 5 stars4.5/5 (5)