You might also like
- Immunology Unveiled: A Comprehensive Journey through the Human Immune System: Guardians of the Body: The Unseen Heroes of ImmunityFrom EverandImmunology Unveiled: A Comprehensive Journey through the Human Immune System: Guardians of the Body: The Unseen Heroes of ImmunityNo ratings yet
- Insect ImmunologyFrom EverandInsect ImmunologyNancy E. BeckageNo ratings yet
- Mini Revision de Inmunidad de La MucosaDocument7 pagesMini Revision de Inmunidad de La MucosaEDGAR FUENMAYOR PIRELANo ratings yet
- A Review On The Interactions Between Gut Microbiota and Innate Immunity of SHDocument10 pagesA Review On The Interactions Between Gut Microbiota and Innate Immunity of SHMarina Tie ShimadaNo ratings yet
- Review: Role of The Microbiota in Immunity and InflammationDocument21 pagesReview: Role of The Microbiota in Immunity and InflammationJULIANA VITORIANo ratings yet
- 1 s2.0 S0952791521001199 MainDocument7 pages1 s2.0 S0952791521001199 MainTatta CamposNo ratings yet
- Ni Hms 276721Document20 pagesNi Hms 276721Jayanthi Mandasari NasutionNo ratings yet
- Fundamental of VaccineDocument7 pagesFundamental of VaccinecarlsonrenovatioNo ratings yet
- 2021 Dawod Et AlDocument10 pages2021 Dawod Et Alpankaj27dataNo ratings yet
- Molecular Mechanisms Used by Salmonella To Evade The Immune SystemDocument35 pagesMolecular Mechanisms Used by Salmonella To Evade The Immune SystemANDREA FERNANDA MUÑOZ NARVAEZNo ratings yet
- The Microbiome and Regulation of Mucosal ImmunityDocument8 pagesThe Microbiome and Regulation of Mucosal ImmunityLevente BalázsNo ratings yet
- Inmune 19Document9 pagesInmune 19Pablo Pinaya SotomayorNo ratings yet
- Immuno 13Document4 pagesImmuno 13thtu.sint.swuNo ratings yet
- Host Microbiomes and Their Interactions With Immune SystemDocument9 pagesHost Microbiomes and Their Interactions With Immune SystemGerald John PazNo ratings yet
- Gut Immune Homeostasis The Immunomodulatory Role of Bacillus Clausii From Basic To Clinical EvidenceDocument14 pagesGut Immune Homeostasis The Immunomodulatory Role of Bacillus Clausii From Basic To Clinical EvidenceBruno GonçalvesNo ratings yet
- Role of Microbiota in Immunity and InflammationDocument12 pagesRole of Microbiota in Immunity and InflammationGerald John PazNo ratings yet
- Immunity of The Greater Wax Moth Galleria Mellonella: ReviewDocument16 pagesImmunity of The Greater Wax Moth Galleria Mellonella: Reviewyaco moturraNo ratings yet
- Epithelial Cell Sense BacteriaDocument5 pagesEpithelial Cell Sense Bacteriau77No ratings yet
- Part 5Document385 pagesPart 5mv.daniborgesNo ratings yet
- Intestinal Epithelial Cells Regulators of Barrier Function and Immune HomeostasisDocument13 pagesIntestinal Epithelial Cells Regulators of Barrier Function and Immune HomeostasisCristian Gutiérrez VeraNo ratings yet
- Immunology KubyDocument24 pagesImmunology KubySayanta Bera100% (1)
- Li LBM 2 SGD 7 Maulana Dika Dinata 31102200075Document4 pagesLi LBM 2 SGD 7 Maulana Dika Dinata 31102200075Devent YanuarNo ratings yet
- 2-3 AnatomicalDocument11 pages2-3 AnatomicalIbnu ZakiNo ratings yet
- Regional Immunity Rakan Fayez El-Hamad: 6 - SecondDocument9 pagesRegional Immunity Rakan Fayez El-Hamad: 6 - SecondInnocent L NdambakuwaNo ratings yet
- Articulo Cientifico de InmunologiaDocument9 pagesArticulo Cientifico de InmunologiaAlejandra RomànNo ratings yet
- Recent Insight Into Autophagy and Immunity at The Maternal-Fetal InterfaceDocument10 pagesRecent Insight Into Autophagy and Immunity at The Maternal-Fetal InterfaceWaldo IzyNo ratings yet
- Evasion InmuneDocument8 pagesEvasion Inmunenydiacastillom2268No ratings yet
- The Vaginal Microenvironment: The Physiologic Role of LactobacilliDocument11 pagesThe Vaginal Microenvironment: The Physiologic Role of Lactobacillishuvo barmanNo ratings yet
- Potential Role of Nutrients On Immunity: Mini ReviewDocument11 pagesPotential Role of Nutrients On Immunity: Mini ReviewEvi NopiantiiNo ratings yet
- Ibrahim Et Al., 2016 PDFDocument11 pagesIbrahim Et Al., 2016 PDFDiba Eka DiputriNo ratings yet
- Sistem Imun UptodateDocument21 pagesSistem Imun UptodateAndreea AlexandruNo ratings yet
- Basic ImmunologyDocument29 pagesBasic ImmunologyHema Duddukuri0% (1)
- Poultry Vaccine LongDocument31 pagesPoultry Vaccine LongLeo HiddenValleyNo ratings yet
- Review: Bacterial Interactions With The Host EpitheliumDocument16 pagesReview: Bacterial Interactions With The Host Epitheliumshaznay delacruzNo ratings yet
- INTESTINO FELIZ, ANIMAL SALUDABLE EsDocument22 pagesINTESTINO FELIZ, ANIMAL SALUDABLE EsurielNo ratings yet
- Club 04Document13 pagesClub 04Muñoz Cerón Smiht YhoissNo ratings yet
- The Impact of Perinatal Immune DevelopmentDocument15 pagesThe Impact of Perinatal Immune DevelopmentAndrés Nicolás Beltrán SeguraNo ratings yet
- (Beri, 2018)Document6 pages(Beri, 2018)Ahmad Bayquni BayquniNo ratings yet
- Immunoglobulins, Mucosal Immunity and Vaccination in Teleost FishDocument14 pagesImmunoglobulins, Mucosal Immunity and Vaccination in Teleost FishBayuNo ratings yet
- The Mucus and Mucins of The Goblet Cells and EnterocytesDocument23 pagesThe Mucus and Mucins of The Goblet Cells and EnterocytesMacario OtsögaNo ratings yet
- Microbiota Intestinal e Inmunidad. Review.2020Document15 pagesMicrobiota Intestinal e Inmunidad. Review.2020hacek357No ratings yet
- Accepted Manuscript: Virus ResearchDocument31 pagesAccepted Manuscript: Virus ResearchNurfitrianti ArfahNo ratings yet
- Immunomodulatory Effects of Probiotics in The Intestinal TractDocument19 pagesImmunomodulatory Effects of Probiotics in The Intestinal TractputrinaraheswariNo ratings yet
- Skin Immunity and Its Dysregulation in Psoriasis: Cell CycleDocument10 pagesSkin Immunity and Its Dysregulation in Psoriasis: Cell CycleJesus MoralesNo ratings yet
- Airway Epithelial Cells: Barrier and Much More: Review ArticleDocument7 pagesAirway Epithelial Cells: Barrier and Much More: Review Articleloup rapideNo ratings yet
- The Basis For The Immunoregulatory Role of Macrophages and Other Accessory Cells.Document7 pagesThe Basis For The Immunoregulatory Role of Macrophages and Other Accessory Cells.Cristian Gutiérrez VeraNo ratings yet
- 1 s2.0 S0065128123001228 MainDocument2 pages1 s2.0 S0065128123001228 MainAinaNo ratings yet
- Basic ImmunologyDocument9 pagesBasic Immunologyarsyiadlina100% (1)
- 1 s2.0 S0753332223002147 MainDocument16 pages1 s2.0 S0753332223002147 MainDr. Iftikhar AliNo ratings yet
- Fukuda 2014 - 2Document5 pagesFukuda 2014 - 2imeldafitriNo ratings yet
- Ijms 24 01526 v2Document25 pagesIjms 24 01526 v2della.ps2310No ratings yet
- Nnate Immune Memory The Missing Piece of The Immunological ResponseDocument12 pagesNnate Immune Memory The Missing Piece of The Immunological ResponseJoss García SalvadorNo ratings yet
- Invited Review: Microbiota of The Bovine Udder: Contributing FactorsDocument21 pagesInvited Review: Microbiota of The Bovine Udder: Contributing FactorsLina Salazar TorresNo ratings yet
- Arsi University: Review On Mucosal Immune ResponseDocument17 pagesArsi University: Review On Mucosal Immune Responsehngdf bfbnNo ratings yet
- The Immune System - An Integrative Perspective - Natural Medicine JournalDocument11 pagesThe Immune System - An Integrative Perspective - Natural Medicine JournalMonaNo ratings yet
- Ojsadmin, 1491Document4 pagesOjsadmin, 1491Cristian PeñaNo ratings yet
- K Upper 2004Document12 pagesK Upper 2004Võ Thành DuyNo ratings yet
- Artis 2008Document10 pagesArtis 2008contramondum1232039No ratings yet
- Cellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsFrom EverandCellular and Molecular Mechanisms of Inflammation: Receptors of Inflammatory Cells: Structure—Function RelationshipsCharles G. CochraneNo ratings yet
- What Is Peripheral Arterial Disease (PAD) ?: Blood VesselsDocument6 pagesWhat Is Peripheral Arterial Disease (PAD) ?: Blood VesselsmainehoonaNo ratings yet
- Infective EndocarditisDocument4 pagesInfective EndocarditismainehoonaNo ratings yet
- MyoclonusDocument4 pagesMyoclonusmainehoonaNo ratings yet
- Clinical Features of ParaperesisDocument31 pagesClinical Features of ParaperesismainehoonaNo ratings yet
- EXtra Info MDDocument62 pagesEXtra Info MDmainehoonaNo ratings yet
- Opthalmalogy CaseDocument11 pagesOpthalmalogy CasemainehoonaNo ratings yet
- Malignant Parotid TumoursDocument11 pagesMalignant Parotid TumoursmainehoonaNo ratings yet
- Peripheral Artery DiseaseDocument8 pagesPeripheral Artery DiseasemainehoonaNo ratings yet
- ChoreaDocument6 pagesChoreamainehoonaNo ratings yet
- Anaesthesia PreopDocument5 pagesAnaesthesia PreopmainehoonaNo ratings yet
- Movement Disorders Include:: and Lower Lip)Document7 pagesMovement Disorders Include:: and Lower Lip)mainehoonaNo ratings yet
- POPDocument42 pagesPOPmainehoonaNo ratings yet
- Huntington's Disease: Liver Disease Dystonia VomitingDocument5 pagesHuntington's Disease: Liver Disease Dystonia VomitingmainehoonaNo ratings yet
- Risk Factors For DiabetesDocument2 pagesRisk Factors For Diabetesmainehoona100% (1)
- Diabetes InsipidusDocument2 pagesDiabetes InsipidusmainehoonaNo ratings yet
- HypertensionDocument2 pagesHypertensionmainehoonaNo ratings yet
- The Family HouseDocument8 pagesThe Family HousemainehoonaNo ratings yet
- Shirazi Salad: IngredientsDocument2 pagesShirazi Salad: IngredientsmainehoonaNo ratings yet
- Hyderabad Biriyani RecipeDocument2 pagesHyderabad Biriyani RecipemainehoonaNo ratings yet
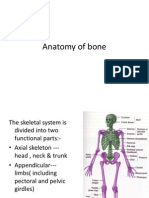
- Anatomy of BoneDocument44 pagesAnatomy of BonemainehoonaNo ratings yet
- Blood Supply of Long BonesDocument4 pagesBlood Supply of Long BonesmainehoonaNo ratings yet
- Turn On Your Favourite Film and Relax in Front of The TV With The Ultimate Caramel Popcorn!Document2 pagesTurn On Your Favourite Film and Relax in Front of The TV With The Ultimate Caramel Popcorn!mainehoonaNo ratings yet
- POPDocument42 pagesPOPmainehoonaNo ratings yet
- PubertyDocument1 pagePubertymainehoonaNo ratings yet
- Human Papilloma Virus (HPV)Document3 pagesHuman Papilloma Virus (HPV)mainehoona100% (1)
- ObesityDocument8 pagesObesitymainehoonaNo ratings yet
- Chocolate-Orange Carrot Cake: IngredientsDocument2 pagesChocolate-Orange Carrot Cake: IngredientsmainehoonaNo ratings yet
- MRI in StagingDocument25 pagesMRI in StagingmainehoonaNo ratings yet
- Cancer Prevention: 7 Tips To Reduce Your RiskDocument3 pagesCancer Prevention: 7 Tips To Reduce Your RiskmainehoonaNo ratings yet
- ObesityDocument8 pagesObesitymainehoonaNo ratings yet
- CCC No: Ministry of Health Effective 01 October 2016 Viral Load Requisition FormDocument1 pageCCC No: Ministry of Health Effective 01 October 2016 Viral Load Requisition FormMigori ArtNo ratings yet
- Which Birds Are Considered Pests?Document4 pagesWhich Birds Are Considered Pests?Liddle RascalsNo ratings yet
- Assessment of Cardiovascular SystemDocument41 pagesAssessment of Cardiovascular Systembicariato100% (7)
- Breast ExamDocument4 pagesBreast Exammoony_sasNo ratings yet
- Ansci Lab 4Document4 pagesAnsci Lab 4Aleah TyNo ratings yet
- Polishing and TrimmingDocument9 pagesPolishing and TrimmingKimberly RodilNo ratings yet
- The Baiting of Wild Animals in RussiaDocument36 pagesThe Baiting of Wild Animals in RussiaZarya LaikaNo ratings yet
- The Horse and Cattle ShowDocument2 pagesThe Horse and Cattle ShowmchinoyNo ratings yet
- AR Code Pokemon White 2Document249 pagesAR Code Pokemon White 2Sir VathNo ratings yet
- Msapila PoutryDocument13 pagesMsapila PoutryEdgar Mugarura100% (2)
- Myo Pro Eng WebDocument12 pagesMyo Pro Eng WebXavier AltamiranoNo ratings yet
- Po Baruna Vasutama Ud Pepito Tuban 10092019Document1 pagePo Baruna Vasutama Ud Pepito Tuban 10092019HizkiaNo ratings yet
- Diseases Management in Aquaculture For Sustainable Food Production PDFDocument34 pagesDiseases Management in Aquaculture For Sustainable Food Production PDFHafiz JamaludinNo ratings yet
- Grand Canyon South Rim Pocket MapDocument2 pagesGrand Canyon South Rim Pocket MapkkkkhineNo ratings yet
- Strength and Conditioning Biological Principles AnDocument8 pagesStrength and Conditioning Biological Principles AnEmanuele DanieleNo ratings yet
- Diskrip Text 2Document11 pagesDiskrip Text 2AdirajaBannederorBallisticNo ratings yet
- Ch2 IS3 TestDocument12 pagesCh2 IS3 TestJohn Smith100% (1)
- The Rights of Animals: A Very Short PrimerDocument15 pagesThe Rights of Animals: A Very Short PrimerajkochanowiczNo ratings yet
- Bomba A Et Al 2002 Probiotics and PUFAsDocument5 pagesBomba A Et Al 2002 Probiotics and PUFAsSandip PatilNo ratings yet
- Anatomy and Physiology of EarDocument11 pagesAnatomy and Physiology of EarAriguna WijayaNo ratings yet
- Newsletter End of Term 1 Grade 1-10Document15 pagesNewsletter End of Term 1 Grade 1-10api-245274300No ratings yet
- Structure of Brain and Its FunctionDocument12 pagesStructure of Brain and Its FunctionDr.Shahana banuNo ratings yet
- SUPPLIERS SPEED DIAL-UpdatedDocument2 pagesSUPPLIERS SPEED DIAL-Updatedlyn7387No ratings yet
- Saratoga: Chileno Entered For Main Track OnlyDocument33 pagesSaratoga: Chileno Entered For Main Track Onlyjohn qNo ratings yet
- Rehaan RamjithDocument2 pagesRehaan RamjithRehaan RamjithNo ratings yet
- ElvesDocument3 pagesElvesCesar ManNo ratings yet
- TB1 Chapter 11 - Multiple ChoiceDocument57 pagesTB1 Chapter 11 - Multiple ChoiceMonicaNo ratings yet
- Mahendravarman III: ArdhamantapaDocument3 pagesMahendravarman III: ArdhamantapaSeema SharmaNo ratings yet
- Practical Research 1 Group2Document12 pagesPractical Research 1 Group2Ron Marc Maranan78% (9)
- Practical Archaeology Training Course - 2004Document34 pagesPractical Archaeology Training Course - 2004Wessex ArchaeologyNo ratings yet