You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Tecnica Sandiwch PDFDocument10 pagesTecnica Sandiwch PDFEddy MazariegosNo ratings yet
- Use of Contact Testing in The Characterization and Design of All-Ceramic Crownlike Layer Structures - A ReviewDocument16 pagesUse of Contact Testing in The Characterization and Design of All-Ceramic Crownlike Layer Structures - A ReviewEddy MazariegosNo ratings yet
- Photographs TipsDocument1 pagePhotographs TipsEddy MazariegosNo ratings yet
- The Importance of Width and Length Ratios in Anterior Pemanent Dentition PDFDocument11 pagesThe Importance of Width and Length Ratios in Anterior Pemanent Dentition PDFEddy Mazariegos100% (1)
- Cosmetic Klaff PDFDocument11 pagesCosmetic Klaff PDFEddy MazariegosNo ratings yet
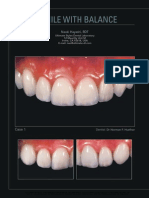
- Smile With Balance PDFDocument9 pagesSmile With Balance PDFEddy MazariegosNo ratings yet
- Using Silicone Impression Material and Acrylic Resin To Fabricate Remount Casts For Removable Partial Dentures PDFDocument5 pagesUsing Silicone Impression Material and Acrylic Resin To Fabricate Remount Casts For Removable Partial Dentures PDFEddy MazariegosNo ratings yet
- Natural Head Position in Standing Subjects PDFDocument17 pagesNatural Head Position in Standing Subjects PDFEddy MazariegosNo ratings yet
- Libman, WJ. Load Fatigue of Teeth Restored With Cast Posts and Cores and Complete CrownsDocument8 pagesLibman, WJ. Load Fatigue of Teeth Restored With Cast Posts and Cores and Complete CrownsEddy Mazariegos100% (1)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- 5.casts and MoldsDocument30 pages5.casts and MoldsKatrina PonceNo ratings yet
- Nasal ObstructionDocument6 pagesNasal Obstructionjoal510No ratings yet
- Basic Position Where Most Exercises BeginDocument9 pagesBasic Position Where Most Exercises Beginantonettedgreat67% (3)
- 2 NeuroanatomyDocument60 pages2 NeuroanatomyMeirina KhairatNo ratings yet
- Introduction 2019 Seminars-in-OrthodonticsDocument2 pagesIntroduction 2019 Seminars-in-OrthodonticsOmy J. CruzNo ratings yet
- Abbé - EstlanderDocument4 pagesAbbé - EstlanderKoretta Esmeral AtehortúaNo ratings yet
- Thyroid and Parathyroid GlandsDocument5 pagesThyroid and Parathyroid GlandsChristopher PhilipsNo ratings yet
- Musculoskeletal Ultrasound Knee ProtocolDocument8 pagesMusculoskeletal Ultrasound Knee ProtocolSabahatAli100% (1)
- Class 15 - Extra Points Part 2: Points M-CA-18 Zi Gong XueDocument6 pagesClass 15 - Extra Points Part 2: Points M-CA-18 Zi Gong XuekillimandjarogeeettNo ratings yet
- Polyvagal TheoryDocument24 pagesPolyvagal Theorymkrnhr100% (7)
- Prosthetics and OrthoticsDocument53 pagesProsthetics and OrthoticsAkbar AziziNo ratings yet
- Shoulder Injuries in Baseball PlayersDocument7 pagesShoulder Injuries in Baseball PlayersMahayu Firsty RamadhaniNo ratings yet
- The Small Intestine Channel of Hand TaiyangDocument5 pagesThe Small Intestine Channel of Hand Taiyanga_steri_x2000No ratings yet
- The Importance of Grip StrengthDocument8 pagesThe Importance of Grip StrengthJosh MyersNo ratings yet
- Oral MucosaDocument4 pagesOral MucosaKathleen Cay Medrano PanopioNo ratings yet
- ManualDocument95 pagesManualcristina_c_43No ratings yet
- Hyperthyroidism: Annals of Internal Medicine July 2012Document17 pagesHyperthyroidism: Annals of Internal Medicine July 2012Deepa RajNo ratings yet
- Prometric Test Part OneDocument167 pagesPrometric Test Part OneMuhammad FahmyNo ratings yet
- BSC 6 Sem General Mammalian Physiology 2 Zoology Summer 2018Document1 pageBSC 6 Sem General Mammalian Physiology 2 Zoology Summer 2018Ujjwala KumbhalkarNo ratings yet
- Understanding The Physical Changes of Puberty: Maria R. de Guzman, Extension Adolescent SpecialistDocument2 pagesUnderstanding The Physical Changes of Puberty: Maria R. de Guzman, Extension Adolescent SpecialistBrett DawsonNo ratings yet
- STBDFSN21Document2 pagesSTBDFSN21Planet BerryNo ratings yet
- Trixie - Anatomy and Physiology (Brain Tumor)Document4 pagesTrixie - Anatomy and Physiology (Brain Tumor)Trixie Marie Sabile AbdullaNo ratings yet
- Group 1 Potd Part 2Document1 pageGroup 1 Potd Part 2Regina CabelisNo ratings yet
- SRB Bedside ClinicDocument592 pagesSRB Bedside ClinicAbinesh Kumar100% (1)
- Materi Dan Latihan Soal Part of BodyDocument2 pagesMateri Dan Latihan Soal Part of BodySeferagic FatihNo ratings yet
- Zach Roys Powerbuilding Program For Men MASTERDocument51 pagesZach Roys Powerbuilding Program For Men MASTERStandaardStanNo ratings yet
- 1.2.2.a SkeletonHunt 2016 - 17 No ConDocument4 pages1.2.2.a SkeletonHunt 2016 - 17 No ConjayNo ratings yet
- CASE REPORT Hip DislocationDocument42 pagesCASE REPORT Hip DislocationvereriNo ratings yet
- Ophthalmology CasesDocument16 pagesOphthalmology CasesDr HappyNo ratings yet
- Workout Exercise Progression Sets Reps Rest (Mins) Notes: Level 2 - Week 1Document8 pagesWorkout Exercise Progression Sets Reps Rest (Mins) Notes: Level 2 - Week 1Michael ANo ratings yet