You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Cognitive Psychology and Its Implications 8th Edition Anderson Test BankDocument20 pagesCognitive Psychology and Its Implications 8th Edition Anderson Test BankJacobFoxgipbn100% (18)
- AP Psych Prep 3 - Biological Psychology (Part II)Document36 pagesAP Psych Prep 3 - Biological Psychology (Part II)dms727No ratings yet
- AP Psych CH 2 Study Guide-Myers 8th EditionDocument7 pagesAP Psych CH 2 Study Guide-Myers 8th Editionlcbell92No ratings yet
- A Glossary: of Key Brain Science TermsDocument24 pagesA Glossary: of Key Brain Science TermscadulemosNo ratings yet
- Script For Teaching Children About MindsetDocument12 pagesScript For Teaching Children About MindsetAnnisa Aulia FitriNo ratings yet
- Neuroimaging Studies of Mental Rotation: A Meta-Analysis and ReviewDocument19 pagesNeuroimaging Studies of Mental Rotation: A Meta-Analysis and ReviewTASYA SALSABILA ISKANDAR 2018No ratings yet
- AconceptualproposalDocument11 pagesAconceptualproposalVÍCTOR MANUEL NEIRA RUBIONo ratings yet
- SDMPSI TutorialDocument18 pagesSDMPSI TutorialStachen SharpenerNo ratings yet
- A Complex Relation Between Depression and Multiple Sclerosis: A Descriptive ReviewDocument8 pagesA Complex Relation Between Depression and Multiple Sclerosis: A Descriptive ReviewGilvan ValdevinoNo ratings yet
- Memory Foundations and Applications 3rd Edition Schwartz Test BankDocument35 pagesMemory Foundations and Applications 3rd Edition Schwartz Test Bankxaviadaniela75xv5100% (12)
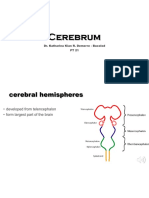
- Cerebrum: Dr. Katharina Kian R. Demerre - Bacolod PT 21Document111 pagesCerebrum: Dr. Katharina Kian R. Demerre - Bacolod PT 21Hephzibah JaporNo ratings yet
- Morphometric Study of Frontal Horn of Lateral Ventricles of The Brain by Computed Tomography in Western Up PopulationDocument6 pagesMorphometric Study of Frontal Horn of Lateral Ventricles of The Brain by Computed Tomography in Western Up Populationrhegi isdiara FairuzNo ratings yet
- Methadone Effects On Frontal Brain Lobe Based EEG-P300 Waves in Drug Rehabilitation PatientsDocument7 pagesMethadone Effects On Frontal Brain Lobe Based EEG-P300 Waves in Drug Rehabilitation PatientsGilbert SihombingNo ratings yet
- Complementary Therapies in Medicine: Yun-Ah Oh, Sin-Ae Park, Byung-Eun AhnDocument5 pagesComplementary Therapies in Medicine: Yun-Ah Oh, Sin-Ae Park, Byung-Eun AhnNelloMasielloNo ratings yet
- Bloa Study BookletDocument65 pagesBloa Study BookletTimedream NightNo ratings yet
- Paul Broca's Historic CasesDocument10 pagesPaul Broca's Historic CasesJaime VerdugoNo ratings yet
- Brune 2006Document14 pagesBrune 2006Dr. Kaushal Kishor SharmaNo ratings yet
- Behavioural and Brain Responses Related To Internet Search and MemoryDocument9 pagesBehavioural and Brain Responses Related To Internet Search and MemoryMinh PhamNo ratings yet
- The Neuroscience of Language. On Brain Circuits of Words and Serial Order by Friedemann PulvermüllerDocument332 pagesThe Neuroscience of Language. On Brain Circuits of Words and Serial Order by Friedemann PulvermüllerMauricio Escobar CastañedaNo ratings yet
- Cases Files NeuroDocument214 pagesCases Files NeurovgdfhgdfgNo ratings yet
- Stroke: Physiotherapy Treatment ApproachesDocument92 pagesStroke: Physiotherapy Treatment ApproachesHimani Kaushik100% (1)
- Adult Development and Aging Canadian 1st Edition Cavanaugh Test BankDocument23 pagesAdult Development and Aging Canadian 1st Edition Cavanaugh Test Bankbryanharrismpqrsfbokn100% (12)
- (Psychology of Emotions Motivations and Actions Series.) Steinmetz, Katherine Mickley - Kensinger, Elizabeth A. - Emotion's Effects On Attention and Memory - Relevance To Posttraumatic Stress DisorderDocument90 pages(Psychology of Emotions Motivations and Actions Series.) Steinmetz, Katherine Mickley - Kensinger, Elizabeth A. - Emotion's Effects On Attention and Memory - Relevance To Posttraumatic Stress DisorderLebron FakesNo ratings yet
- 2-Brain, Evolution ofDocument8 pages2-Brain, Evolution ofsantiagoNo ratings yet
- Ebook Fitzgeralds Clinical Neuroanatomy and Neuroscience PDF Full Chapter PDFDocument67 pagesEbook Fitzgeralds Clinical Neuroanatomy and Neuroscience PDF Full Chapter PDFcharles.cunningham811100% (26)
- Vocabulary Brain 1Document6 pagesVocabulary Brain 1Şterbeţ RuxandraNo ratings yet
- MeditationDocument114 pagesMeditationdBiswas2011100% (2)
- Aaron Hernandez Case Study 1.2Document3 pagesAaron Hernandez Case Study 1.2shevarkNo ratings yet
- Tourville - Guenther 2013 Actualizacion DIVA Model PDFDocument27 pagesTourville - Guenther 2013 Actualizacion DIVA Model PDFDaniel Gaete OsorioNo ratings yet
- Corteza Prefrontal Memoria y AtencionDocument5 pagesCorteza Prefrontal Memoria y AtencionNicoleCalvillanNo ratings yet