You might also like
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Intestinal Malrotation and VolvulusDocument14 pagesIntestinal Malrotation and VolvulusSaf Tanggo Diampuan100% (1)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- AKUH Kampala Hospital-Press Release - Final - 17 December 2015Document3 pagesAKUH Kampala Hospital-Press Release - Final - 17 December 2015Robert OkandaNo ratings yet
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Ummer Roject Resentation ON Herbal Healthcare Industry in IndiaDocument14 pagesUmmer Roject Resentation ON Herbal Healthcare Industry in IndiaSaurabh RustagiNo ratings yet
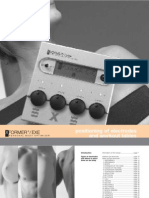
- User Manual For Applications Xformer Exe EnglishLightDocument51 pagesUser Manual For Applications Xformer Exe EnglishLightNunoClaudinoNo ratings yet
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- CeftriaxoneDocument2 pagesCeftriaxoneFlora Angeli PastoresNo ratings yet
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Nephrolithiasis PDFDocument21 pagesNephrolithiasis PDFAulia AlmaNo ratings yet
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Curriculum Vitae (CV)Document4 pagesCurriculum Vitae (CV)wubeshetNo ratings yet
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Biology SPMDocument17 pagesBiology SPMbloptra18No ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- 2 Articles From Oui Magazine (Feat. Robert Anton Wilson)Document16 pages2 Articles From Oui Magazine (Feat. Robert Anton Wilson)Jakob AndradeNo ratings yet
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Cutaneous Candidiasis - An Evidence-Based Review of Topical and Systemic Treatments To Inform Clinical PracticeDocument11 pagesCutaneous Candidiasis - An Evidence-Based Review of Topical and Systemic Treatments To Inform Clinical PracticeRose ParkNo ratings yet
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Nabh Entry LevelDocument64 pagesNabh Entry LevelRenuka MuruganNo ratings yet
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- FilariasisDocument5 pagesFilariasisSathish KumarNo ratings yet
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Growth RotationDocument57 pagesGrowth RotationAtul Koundel100% (1)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- September 2013 Exam QuestionsDocument13 pagesSeptember 2013 Exam QuestionsooiziungieNo ratings yet
- PackagingDocument29 pagesPackagingSagar TummaNo ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- PIL 18047 LatestDocument2 pagesPIL 18047 LatestWendy EscalanteNo ratings yet
- Cardiac - Anatomy of The HeartDocument9 pagesCardiac - Anatomy of The HeartRiya ThoratNo ratings yet
- Psychiatry MCQDocument12 pagesPsychiatry MCQMonika Joseph100% (8)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Funny StoriesDocument5 pagesFunny Storiesamin jamalNo ratings yet
- Acticide Bac 50 M MSDS PDFDocument10 pagesActicide Bac 50 M MSDS PDFmeNo ratings yet
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Effects of Skin Bleach in SocietyDocument8 pagesThe Effects of Skin Bleach in SocietySalma BahNo ratings yet
- 12.4 Female Reproductive DiseasesDocument20 pages12.4 Female Reproductive Diseasessanat kr pratiharNo ratings yet
- Sport Psychology ToolsDocument169 pagesSport Psychology Toolsfeldexxor100% (5)
- WC500059147 PDFDocument140 pagesWC500059147 PDFAnaStankovićNo ratings yet
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Pediatric Unfolding Case StudyDocument4 pagesPediatric Unfolding Case StudyJessica100% (1)
- John O. Agwunobi MD Mba MPHDocument1 pageJohn O. Agwunobi MD Mba MPHStalyn DíazNo ratings yet
- Hostel Prospectus BookletDocument12 pagesHostel Prospectus BookletsansharmajsNo ratings yet
- Orthodontics!!Document9 pagesOrthodontics!!Ayesha AwanNo ratings yet
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- AutismAdvisoryPanelReport 2019Document64 pagesAutismAdvisoryPanelReport 2019CityNewsTorontoNo ratings yet
- Canine and Feline Mega EsophagusDocument10 pagesCanine and Feline Mega Esophagustaner_soysurenNo ratings yet
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)