You might also like
- Proud Foot 2002Document12 pagesProud Foot 2002Mumun Hal MuratNo ratings yet
- Christian Kambach, Stefan Walke and Kiyoshi NagaiDocument9 pagesChristian Kambach, Stefan Walke and Kiyoshi NagaiDivya NarayanNo ratings yet
- New Functions of Small Nucleolar RNAsDocument13 pagesNew Functions of Small Nucleolar RNAsLeslie ARaNo ratings yet
- Transport Dependent Proteolysis of SREBP CellDocument10 pagesTransport Dependent Proteolysis of SREBP CellGabriel RomoNo ratings yet
- Post Transcriptional ModificationDocument31 pagesPost Transcriptional ModificationranasiddharthNo ratings yet
- Spliceosome Structure and Function PDFDocument23 pagesSpliceosome Structure and Function PDFfroywanNo ratings yet
- Pombe, Regulates Spore Wall Formation: Mug28, A Meiosis-Specific Protein of SchizosaccharomycesDocument13 pagesPombe, Regulates Spore Wall Formation: Mug28, A Meiosis-Specific Protein of SchizosaccharomycesleongchinrickNo ratings yet
- 002 - 2018 - Splicing Mutations in Human Genetic Disorders - Examples Detection and ConfirmationDocument16 pages002 - 2018 - Splicing Mutations in Human Genetic Disorders - Examples Detection and ConfirmationAndrea Bermúdez QuinteroNo ratings yet
- RNA-2008-Dávila López-1-10Document11 pagesRNA-2008-Dávila López-1-10akNo ratings yet
- Mintz 1999Document13 pagesMintz 1999Laura MirceaNo ratings yet
- tmp7F3E TMPDocument6 pagestmp7F3E TMPFrontiersNo ratings yet
- tmp83FE TMPDocument11 pagestmp83FE TMPFrontiersNo ratings yet
- Mammalian NET-seq Analysis Defines Nascent RNA Profiles and Associated RNA Processing Genome-WideDocument16 pagesMammalian NET-seq Analysis Defines Nascent RNA Profiles and Associated RNA Processing Genome-WideLorenzo AmpilNo ratings yet
- RNA SplicingDocument8 pagesRNA SplicingDivya NarayanNo ratings yet
- Resna N K MicrobiologyDocument54 pagesResna N K MicrobiologyResna N K ResiNo ratings yet
- MBC 7 10 1559-2Document14 pagesMBC 7 10 1559-2Laura MirceaNo ratings yet
- Different Types of RNA Molecule and Their Functional Role in CellDocument8 pagesDifferent Types of RNA Molecule and Their Functional Role in CellNoor-E-Khadiza ShamaNo ratings yet
- Analisis Funcional de La Familia de NRAMPDocument9 pagesAnalisis Funcional de La Familia de NRAMPQuimico Inmunologia GeneticaNo ratings yet
- Tau Filaments in Neurodegenerative Diseases Goedert-2018-FEBS - LettersDocument30 pagesTau Filaments in Neurodegenerative Diseases Goedert-2018-FEBS - LettersElena Martínez CelisNo ratings yet
- L5-6 Translation in ProkaryotesDocument17 pagesL5-6 Translation in ProkaryotesArchana BhartiNo ratings yet
- Teratozoospermia in Mice Lacking The Transition Protein 2 (Tnp2)Document8 pagesTeratozoospermia in Mice Lacking The Transition Protein 2 (Tnp2)BerlinNo ratings yet
- Eukaryotic Gene Regulation: Co-Transcriptional and Post-Transcriptional Modifications of Pre Messenger RNA - IDocument28 pagesEukaryotic Gene Regulation: Co-Transcriptional and Post-Transcriptional Modifications of Pre Messenger RNA - Ismart ptmNo ratings yet
- Articulo 9 CovidDocument9 pagesArticulo 9 CovidJuan OrlandoniNo ratings yet
- 10 23 2006 MCB RNA Processing ChengDocument34 pages10 23 2006 MCB RNA Processing Chengapi-3696530No ratings yet
- RIP-seq Analysis of Eukaryotic SM Proteins Identifies Three Major Categories of Sm-Containing RibonucleoproteinsDocument23 pagesRIP-seq Analysis of Eukaryotic SM Proteins Identifies Three Major Categories of Sm-Containing RibonucleoproteinsYunyang LiNo ratings yet
- Neurosci Lett 2009Document5 pagesNeurosci Lett 2009wangyu81No ratings yet
- Rna Processing: M.Prasad Naidu MSC Medical Biochemistry, PH.DDocument33 pagesRna Processing: M.Prasad Naidu MSC Medical Biochemistry, PH.DDr. M. Prasad NaiduNo ratings yet
- Co-Expression and Co-Purification of Archaeal and Eukaryal Box C/D RnpsDocument8 pagesCo-Expression and Co-Purification of Archaeal and Eukaryal Box C/D RnpsvirhelgueraNo ratings yet
- Splicing - NatureDocument4 pagesSplicing - NatureBrijesh KumarNo ratings yet
- Science Abm4245Document8 pagesScience Abm4245Tanvi GargNo ratings yet
- 2022-04-25 L6 - RNA ProcessingDocument43 pages2022-04-25 L6 - RNA ProcessingTamara ElyasNo ratings yet
- The Caenorhabditis Elegans Schnurri Homolog Sma-9 Mediates Stage-And Cell Type-Specific Responses To DBL-1 BMP-related SignalingDocument12 pagesThe Caenorhabditis Elegans Schnurri Homolog Sma-9 Mediates Stage-And Cell Type-Specific Responses To DBL-1 BMP-related SignalingAziz RosyidNo ratings yet
- Rna Processing EukaryotesDocument33 pagesRna Processing EukaryotesNathanaelNo ratings yet
- 1Document7 pages1hendycoNo ratings yet
- Eukaryotic RNA Processing 28 Jan 2014Document26 pagesEukaryotic RNA Processing 28 Jan 2014madhi ahuNo ratings yet
- #16 Ch21 SplicingDocument31 pages#16 Ch21 SplicingKyle BroflovskiNo ratings yet
- Moroni - Mirko-HS AND LS ALFA4 BETA 2Document14 pagesMoroni - Mirko-HS AND LS ALFA4 BETA 2Omar Hernández GonzálezNo ratings yet
- Vps41 Phosphorylation and The Rab Ypt7 Control The Targeting of The HOPS Complex To Endosome-Vacuole Fusion SitesDocument12 pagesVps41 Phosphorylation and The Rab Ypt7 Control The Targeting of The HOPS Complex To Endosome-Vacuole Fusion SitesMargarita CabreraNo ratings yet
- Transcription in EukaryotesDocument4 pagesTranscription in EukaryotesShaher Bano MirzaNo ratings yet
- 471 - BCH 201-Lecture Note On WebDocument10 pages471 - BCH 201-Lecture Note On WebShaikh SalmanNo ratings yet
- Expression of A Drosophila Circadian ClockDocument12 pagesExpression of A Drosophila Circadian Clocklana valeryNo ratings yet
- Session 10Document10 pagesSession 10VIKRANT SIDANANo ratings yet
- Large-Scale Production of 5 Capped RNA - Fuchs2016Document14 pagesLarge-Scale Production of 5 Capped RNA - Fuchs2016Marcus PlungianskiNo ratings yet
- Spatial Specification of Mammalian Eye Territories by Reciprocal Transcriptional Repression of Pax2 and Pax6Document10 pagesSpatial Specification of Mammalian Eye Territories by Reciprocal Transcriptional Repression of Pax2 and Pax6Carmen Mansilla De LatorreNo ratings yet
- 7.isoform-Specific Regulation of Osteogenic Factors by Polypeptide N-Acetylgalactosaminyltransferases 1 and 4 3Document6 pages7.isoform-Specific Regulation of Osteogenic Factors by Polypeptide N-Acetylgalactosaminyltransferases 1 and 4 3Shawn GaoNo ratings yet
- Characterization of Geranium Chloroplast EF-Tu cDNA: (Pelargonium Graveolens)Document5 pagesCharacterization of Geranium Chloroplast EF-Tu cDNA: (Pelargonium Graveolens)Silvana OliveiraNo ratings yet
- Transcription and TranslationDocument35 pagesTranscription and TranslationMing mingNo ratings yet
- Tau Protein in Neurodegenerative Diseases - A Review: E V I E WDocument10 pagesTau Protein in Neurodegenerative Diseases - A Review: E V I E WyusufNo ratings yet
- SNAP23 Deficiency Causes Severe Brain Dysplasia Through The Loss of Radial Glial Cell Polarity 2021Document28 pagesSNAP23 Deficiency Causes Severe Brain Dysplasia Through The Loss of Radial Glial Cell Polarity 2021Carlos Henrique JardimNo ratings yet
- Poros NuclearesDocument27 pagesPoros NuclearescarlosNo ratings yet
- Protein Sysnthesis 3Document36 pagesProtein Sysnthesis 3Fan SmithNo ratings yet
- Arp2/3 Complex: Arp2/3 Complex Is A Seven-Subunit Protein Complex That Plays ADocument5 pagesArp2/3 Complex: Arp2/3 Complex Is A Seven-Subunit Protein Complex That Plays Afelipe smithNo ratings yet
- Dependence of ORC Silencing Function OnDocument13 pagesDependence of ORC Silencing Function Onkilli999No ratings yet
- The Polycomb Group Protein Yaf2 Regulates The Pluripotency of Embryonic Stem Cells in A Phosphorylation-Dependent MannerDocument13 pagesThe Polycomb Group Protein Yaf2 Regulates The Pluripotency of Embryonic Stem Cells in A Phosphorylation-Dependent MannerSHUMETNo ratings yet
- Processing of Vertebrate Box C/D Small Nucleolar Rnas in Plant CellsDocument7 pagesProcessing of Vertebrate Box C/D Small Nucleolar Rnas in Plant CellsBengt HörbergNo ratings yet
- Tsukushi Functions As An Organizer Inducer by Inhibition of - 2004 - DevelopmenDocument12 pagesTsukushi Functions As An Organizer Inducer by Inhibition of - 2004 - Developmenhalil sahanNo ratings yet
- 2016-Joo-AMPK Phosphorylates NRF2 Ser550Document12 pages2016-Joo-AMPK Phosphorylates NRF2 Ser550HaiNo ratings yet
- LECT-09 Transcription in EukaryotesDocument27 pagesLECT-09 Transcription in EukaryotesFAKHRI HUSAINI NASUTION 1No ratings yet
- Neurotransmission: Proceedings of the Sixth International Congress of PharmacologyFrom EverandNeurotransmission: Proceedings of the Sixth International Congress of PharmacologyLiisa AhteeNo ratings yet
- Biochemical and Pharmacological Roles of Adenosylmethionine and the Central Nervous System: Proceedings of an International Round Table on Adenosylmethionine and the Central Nervous System, Naples, Italy, May 1978From EverandBiochemical and Pharmacological Roles of Adenosylmethionine and the Central Nervous System: Proceedings of an International Round Table on Adenosylmethionine and the Central Nervous System, Naples, Italy, May 1978Vincenzo ZappiaNo ratings yet
- 2.1.2 - PRISMA 2009 ChecklistDocument2 pages2.1.2 - PRISMA 2009 ChecklistFiŗåš ÀßßâşNo ratings yet
- HTN CDCDocument1 pageHTN CDCnemeziennaNo ratings yet
- Licensing Exam ReviewDocument4 pagesLicensing Exam ReviewnemeziennaNo ratings yet
- Cover Letter GuideDocument8 pagesCover Letter Guidenemezienna100% (1)
- 2011 Nuclear Speckles ReviewDocument12 pages2011 Nuclear Speckles ReviewnemeziennaNo ratings yet
- Case Study #2Document2 pagesCase Study #2nemeziennaNo ratings yet
- Kuby Immunology, 7e: Chapter 2: Cells, Organs, and Microenvironments of The Immune SystemDocument37 pagesKuby Immunology, 7e: Chapter 2: Cells, Organs, and Microenvironments of The Immune Systemnemezienna100% (1)
- Jonathan Haidt How Does Moral Judgement WorkDocument7 pagesJonathan Haidt How Does Moral Judgement WorkPaulo LuísNo ratings yet
- Neural Basis of Economic Decision CohenDocument4 pagesNeural Basis of Economic Decision CohennemeziennaNo ratings yet
- Case StudyDocument6 pagesCase StudynemeziennaNo ratings yet
- Stages of LearningDocument24 pagesStages of Learningapi-278802103No ratings yet
- Year - 9 PhotosynthesisDocument10 pagesYear - 9 PhotosynthesisChuanthing TeeNo ratings yet
- Individual Assignment - SwimmingDocument9 pagesIndividual Assignment - Swimminghaafiz5arisNo ratings yet
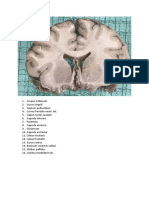
- Brain Slides SEMINAR 1 - 220606 - 142811 - 220606 - 223805Document32 pagesBrain Slides SEMINAR 1 - 220606 - 142811 - 220606 - 223805pang pangNo ratings yet
- Centruroides Sculpturatus (Buthidae) and Diplocentrus Spitzeri (Diplocentridae) - J - ArachnolDocument12 pagesCentruroides Sculpturatus (Buthidae) and Diplocentrus Spitzeri (Diplocentridae) - J - ArachnoladriendrixNo ratings yet
- Journal - A Review of Diabetes MellitusDocument6 pagesJournal - A Review of Diabetes Mellitusifaans16No ratings yet
- Natures Hidden Health MiracleDocument38 pagesNatures Hidden Health MiracleGorneau100% (2)
- Class CHAPTER 9 - Circulatory With AnswersDocument59 pagesClass CHAPTER 9 - Circulatory With AnswersSara VinuyaNo ratings yet
- HypoxiaDocument21 pagesHypoxiaJojo RyelciusNo ratings yet
- 1 2 Ultrastructure of Cells - Students NotesDocument17 pages1 2 Ultrastructure of Cells - Students Notesapi-289234696No ratings yet
- Calf Rearing Practice - ManualDocument33 pagesCalf Rearing Practice - ManualJose DeJesus Bustamante GuerreroNo ratings yet
- Histochemical Uses of HematoxylinDocument11 pagesHistochemical Uses of HematoxylinCarla Victoria Vejar DíazNo ratings yet
- Abdominal IncisionsDocument8 pagesAbdominal IncisionsdkasisNo ratings yet
- Urinary Disorders 2Document19 pagesUrinary Disorders 2ula mdiNo ratings yet
- Acute PeritonitisDocument4 pagesAcute PeritonitisSatrio Tri HadmokoNo ratings yet
- Anatomia de BufalosDocument24 pagesAnatomia de BufalosTrilce ClaroNo ratings yet
- A Primer For The Primary FANZCA ExaminationDocument62 pagesA Primer For The Primary FANZCA ExaminationAnnie100% (1)
- Renal Emergency RevisiDocument103 pagesRenal Emergency Revisidesy f sarahNo ratings yet
- Genetics in Blood BankingDocument47 pagesGenetics in Blood BankingMandy A. Delfin100% (3)
- Nervous System NotesDocument6 pagesNervous System NotesAlex Whitwam100% (3)
- MHD Exam 6 MaterialDocument179 pagesMHD Exam 6 Materialnaexuis5467100% (1)
- 083 41 Final Biologi T4 DLP-30-60Document31 pages083 41 Final Biologi T4 DLP-30-60Farhan DarwisyNo ratings yet
- The Rhesus (RH) Blood Group SystemDocument44 pagesThe Rhesus (RH) Blood Group SystemMarl EstradaNo ratings yet
- 294 Fetal Heart Rate Monitoring. Principles and Interpretation of Cardiotocography LR PDFDocument9 pages294 Fetal Heart Rate Monitoring. Principles and Interpretation of Cardiotocography LR PDFelenNo ratings yet
- Grade 8 Intro To RespirationDocument27 pagesGrade 8 Intro To RespirationPrisha ChhedaNo ratings yet
- Case Study 18Document4 pagesCase Study 18api-340604016100% (1)
- Introduction To TheCourseDocument21 pagesIntroduction To TheCourseCharles SainzNo ratings yet
- L.P On Lambar PunctureDocument16 pagesL.P On Lambar PunctureMary MenuNo ratings yet
- Stress Management Strategies Adopted by Teachers in Public Primary Schools in Obio/Akpor Local Government Area of Rivers State, Nigeria.Document94 pagesStress Management Strategies Adopted by Teachers in Public Primary Schools in Obio/Akpor Local Government Area of Rivers State, Nigeria.sorbariNo ratings yet