You might also like
- AMP-activated Protein Kinase Is Activated As A Consequence J. Biol. Chem.-2008-Gauthier-16514-24Document12 pagesAMP-activated Protein Kinase Is Activated As A Consequence J. Biol. Chem.-2008-Gauthier-16514-24evanconstantine77No ratings yet
- NIH Public Access: Author ManuscriptDocument34 pagesNIH Public Access: Author Manuscriptcamilamadrid01No ratings yet
- AMPK Modulates Tissue and Organismal Aging in A Non-Cell-Autonomous MannerDocument15 pagesAMPK Modulates Tissue and Organismal Aging in A Non-Cell-Autonomous MannerPilar Bravo SalasNo ratings yet
- CDD Is 2013151 ADocument10 pagesCDD Is 2013151 ADalton JLNo ratings yet
- Central Exercise Action Increases The AMPK and mTOR Response To LeptinDocument17 pagesCentral Exercise Action Increases The AMPK and mTOR Response To LeptinMario PérezNo ratings yet
- Appl1 Scaffolds Tak1-Mkk3-p38 Mapk in Adiponectin PathwayDocument9 pagesAppl1 Scaffolds Tak1-Mkk3-p38 Mapk in Adiponectin PathwaypopopioNo ratings yet
- AMPK Phosphorylation Assay KitDocument3 pagesAMPK Phosphorylation Assay KitBioAssay 2015No ratings yet
- Chen2012 Article Α-LipoicAcidRegulatesLipidMetaDocument12 pagesChen2012 Article Α-LipoicAcidRegulatesLipidMetaJoshNo ratings yet
- Glikogen 6Document12 pagesGlikogen 6DZoel FAcHMyNo ratings yet
- Metformin Induces Rab4 Through AMPK and Modulates GLUT4 Translocation in Skeletal Muscle CellsDocument8 pagesMetformin Induces Rab4 Through AMPK and Modulates GLUT4 Translocation in Skeletal Muscle CellsZulvina FaozanudinNo ratings yet
- 1 s2.0 S0753332221009720 MainDocument10 pages1 s2.0 S0753332221009720 MaingabrielaNo ratings yet
- AMPK in Skeletal Muscle Function and Metabolism 2018Document38 pagesAMPK in Skeletal Muscle Function and Metabolism 2018Rita De Cassia Marqueti DuriganNo ratings yet
- Lab Invest 2014131 ADocument12 pagesLab Invest 2014131 AGeorge Sebastian AntonyNo ratings yet
- Inhibition of Rat PC12 Cell Calpain Activity by Glutathione, Oxidized Glutathione and Nitric OxideDocument4 pagesInhibition of Rat PC12 Cell Calpain Activity by Glutathione, Oxidized Glutathione and Nitric OxidePaoloNo ratings yet
- Carling 2004Document7 pagesCarling 2004Jocilene Dantas Torres NascimentoNo ratings yet
- Ampk and The Adaptation To ExerciseDocument3 pagesAmpk and The Adaptation To ExerciseAndrés ChávezNo ratings yet
- Biochimica Et Biophysica Acta: Maayan Barnea, Liyan Haviv, Roee Gutman, Nava Chapnik, Zecharia Madar, Oren FroyDocument11 pagesBiochimica Et Biophysica Acta: Maayan Barnea, Liyan Haviv, Roee Gutman, Nava Chapnik, Zecharia Madar, Oren FroyFernando Caro MenaNo ratings yet
- Ca2calmodulin-Dependent Protein Kinase Kinase Is IDocument11 pagesCa2calmodulin-Dependent Protein Kinase Kinase Is IJoshNo ratings yet
- Ampk Hormonal 2Document11 pagesAmpk Hormonal 2miguel contrerasNo ratings yet
- Ref 17Document10 pagesRef 17Lateecka R KulkarniNo ratings yet
- Pancreatic B-Cell Response To Increased Metabolic Demand and To Pharmacologic Secretagogues Requires EPAC2ADocument12 pagesPancreatic B-Cell Response To Increased Metabolic Demand and To Pharmacologic Secretagogues Requires EPAC2AMunib Ur RehmanNo ratings yet
- 926.full 2Document3 pages926.full 2Nadilla NatashaNo ratings yet
- Lecture 8-CAMP and Cell SignallingDocument7 pagesLecture 8-CAMP and Cell Signallingjosphat nzuvaNo ratings yet
- High-Fat Diet Induces Endothelial Dysfunction Through A Down-Regulation of The Endothelial AMPK-PI3K-Akt-eNOS PathwayDocument13 pagesHigh-Fat Diet Induces Endothelial Dysfunction Through A Down-Regulation of The Endothelial AMPK-PI3K-Akt-eNOS PathwayJorge CamargoNo ratings yet
- FullDocument10 pagesFullWalida FadillahNo ratings yet
- Jurnal ResveratrolDocument10 pagesJurnal ResveratrolAsis FitrianaNo ratings yet
- 2016-Joo-AMPK Phosphorylates NRF2 Ser550Document12 pages2016-Joo-AMPK Phosphorylates NRF2 Ser550HaiNo ratings yet
- AMPK Promove Expressão Gênica de GLUT 4 em Humanos. MCGGE (2008)Document8 pagesAMPK Promove Expressão Gênica de GLUT 4 em Humanos. MCGGE (2008)Anderson Ranieri MassahudNo ratings yet
- Involvement of SIK2/TORC2 Signaling Cascade in The Regulation of Insulin-Induced PGC-1 and UCP-1 Gene Expression in Brown AdipocytesDocument10 pagesInvolvement of SIK2/TORC2 Signaling Cascade in The Regulation of Insulin-Induced PGC-1 and UCP-1 Gene Expression in Brown Adipocytesalejandra soledad alvarado neiraNo ratings yet
- MainDocument8 pagesMainAry MadinaNo ratings yet
- RNABPDocument7 pagesRNABPDr-Dalya ShakirNo ratings yet
- p38 Mitogen-Activated Protein Kinase Is The Central Regulator of Cyclic AMP-Dependent Transcription of The Brown Fat Uncoupling Protein 1 GeneDocument11 pagesp38 Mitogen-Activated Protein Kinase Is The Central Regulator of Cyclic AMP-Dependent Transcription of The Brown Fat Uncoupling Protein 1 GeneAlmir FilsNo ratings yet
- First PaperDocument8 pagesFirst PaperHanan AhmedNo ratings yet
- 2006 Gouda Et Al.Document6 pages2006 Gouda Et Al.Lalitha R GowdaNo ratings yet
- Biochemistry and Biophysics Reports: SciencedirectDocument5 pagesBiochemistry and Biophysics Reports: SciencedirectDAVID VILLANo ratings yet
- CAMPing in The Immune System - News & Announcements - Cayman ChemicalDocument3 pagesCAMPing in The Immune System - News & Announcements - Cayman ChemicalRovin RamphalNo ratings yet
- HMG-CoA Reductase Inhibitors (Statins) Activate Expression of PPARalpha - PPARgamma and ABCA1 in Cultured Gallbladder Epithelial Cells.Document8 pagesHMG-CoA Reductase Inhibitors (Statins) Activate Expression of PPARalpha - PPARgamma and ABCA1 in Cultured Gallbladder Epithelial Cells.Tiago TorresNo ratings yet
- Cold Spring Harb Symp Quant BiolDocument6 pagesCold Spring Harb Symp Quant BioleliNo ratings yet
- tmp90FE TMPDocument10 pagestmp90FE TMPFrontiersNo ratings yet
- Nihms 341890Document18 pagesNihms 341890Alex Fit FitNo ratings yet
- Junior Year Fall - Biochemistry pfk-1 Assay Formal Report Nov 30Document7 pagesJunior Year Fall - Biochemistry pfk-1 Assay Formal Report Nov 30api-651131417No ratings yet
- Froy Et Al 2011Document10 pagesFroy Et Al 2011Roee GutmanNo ratings yet
- HanooDocument8 pagesHanoomr samoNo ratings yet
- Curcumin Effect On ObesityDocument7 pagesCurcumin Effect On ObesityAldo MataNo ratings yet
- Rac1/Pi3K/Pkb-Dependent Caspase-1 Activation Release Through Hmg-Coa Reductase Inhibition Induces Il-1Document12 pagesRac1/Pi3K/Pkb-Dependent Caspase-1 Activation Release Through Hmg-Coa Reductase Inhibition Induces Il-1Josse BouwhuisNo ratings yet
- Research Proposal-Kelsi GelleDocument5 pagesResearch Proposal-Kelsi Gelleapi-619041501No ratings yet
- ANEMIA OLAHRAGA PentingDocument6 pagesANEMIA OLAHRAGA PentingfahmiNo ratings yet
- Dewei Ye, Yudong Wang, Huating Li, Weiping Jia, Kwan Man, Chung Mau Lo, Yu Wang, Karen S.L. Lam, and Aimin XuDocument13 pagesDewei Ye, Yudong Wang, Huating Li, Weiping Jia, Kwan Man, Chung Mau Lo, Yu Wang, Karen S.L. Lam, and Aimin XuzaifNo ratings yet
- Curcumin Modulates The Apolipoprotein B mRNA EditiDocument9 pagesCurcumin Modulates The Apolipoprotein B mRNA EditiLight YagamiNo ratings yet
- Autophagy & TomorigenisisDocument22 pagesAutophagy & TomorigenisisKirk CobainNo ratings yet
- Xiong2011 2Document6 pagesXiong2011 2Alejandro SalazarNo ratings yet
- The Jak/Stat Pathway: A Novel Way To Regulate PI3K ActivityDocument4 pagesThe Jak/Stat Pathway: A Novel Way To Regulate PI3K ActivityskljoleNo ratings yet
- Literature ReviewDocument15 pagesLiterature Reviewapi-619041501No ratings yet
- New Mechanisms of Metformin Action: Focusing On Mitochondria and The GutDocument10 pagesNew Mechanisms of Metformin Action: Focusing On Mitochondria and The GutZulvina FaozanudinNo ratings yet
- Biochemistry: ADP ADP Phosphorylation (Kinase) PDocument7 pagesBiochemistry: ADP ADP Phosphorylation (Kinase) PVivek VinayakumarNo ratings yet
- Diet Effects On MitochondriaDocument26 pagesDiet Effects On MitochondriabenZen100% (1)
- Regulation of Apoa Gene Expression With Acidosis: Requirement For A Transcriptional RepressorDocument15 pagesRegulation of Apoa Gene Expression With Acidosis: Requirement For A Transcriptional RepressorAbdur Rachman Ba'abdullahNo ratings yet
- Peroxisome Proliferator-Activated Receptor Gamma (Ppar C) Regulates Lactase Expression and Activity in The GutDocument11 pagesPeroxisome Proliferator-Activated Receptor Gamma (Ppar C) Regulates Lactase Expression and Activity in The GutrcastacNo ratings yet
- tmp5 TMPDocument8 pagestmp5 TMPFrontiersNo ratings yet
- Cellular Endocrinology in Health and DiseaseFrom EverandCellular Endocrinology in Health and DiseaseAlfredo Ulloa-AguirreNo ratings yet
- 2005 Return To Official Italian First Division Soccer Games Within 90 Days After Anterior Cruciate Ligament Reconstruction A Case ReportDocument15 pages2005 Return To Official Italian First Division Soccer Games Within 90 Days After Anterior Cruciate Ligament Reconstruction A Case ReportFrancisco Antonó Castro WeithNo ratings yet
- 2018 The Truth About Artificial Sweeteners - Are They Good For Diabetics...Document3 pages2018 The Truth About Artificial Sweeteners - Are They Good For Diabetics...Francisco Antonó Castro WeithNo ratings yet
- 2018 Muscle Strength Is A Poor Screening Test For Predicting Lower Extremity Injuries in Professional Male Soccer Players. A 2-Year Prospective Cohort StudyDocument11 pages2018 Muscle Strength Is A Poor Screening Test For Predicting Lower Extremity Injuries in Professional Male Soccer Players. A 2-Year Prospective Cohort StudyFrancisco Antonó Castro WeithNo ratings yet
- 2000 Metabolically Active Components of Fat Free Mass and Resting Energy Expenditure in Nonobese AdultsDocument8 pages2000 Metabolically Active Components of Fat Free Mass and Resting Energy Expenditure in Nonobese AdultsFrancisco Antonó Castro WeithNo ratings yet
- 2018 Association of Efficacy of Resistance Exercise Training With Depressive Symtoms. Meta-Analysis and Meta - Regression Analysis of Randomized Clinical TrialsDocument11 pages2018 Association of Efficacy of Resistance Exercise Training With Depressive Symtoms. Meta-Analysis and Meta - Regression Analysis of Randomized Clinical TrialsFrancisco Antonó Castro WeithNo ratings yet
- 2018 Physical Therapists Forward Deployed On Aircraft Carriers A Retrospective Look at A Decade of ServiceDocument6 pages2018 Physical Therapists Forward Deployed On Aircraft Carriers A Retrospective Look at A Decade of ServiceFrancisco Antonó Castro WeithNo ratings yet
- 2018 A Systematic Review of The Relationship Between Physical Activity and HappinessDocument18 pages2018 A Systematic Review of The Relationship Between Physical Activity and HappinessFrancisco Antonó Castro Weith100% (1)
- 2004 Ability of A New Hop Test To Determine Functional Deficits After Anterior Cruciate Ligament ReconstructionDocument7 pages2004 Ability of A New Hop Test To Determine Functional Deficits After Anterior Cruciate Ligament ReconstructionFrancisco Antonó Castro WeithNo ratings yet
- 2018 The Truth About Artificial Sweeteners - Are They Good For Diabetics...Document3 pages2018 The Truth About Artificial Sweeteners - Are They Good For Diabetics...Francisco Antonó Castro WeithNo ratings yet
- 2018 Football Recovery Strategies. Practical Aspects of Blending Science and RealityDocument8 pages2018 Football Recovery Strategies. Practical Aspects of Blending Science and RealityFrancisco Antonó Castro Weith100% (1)
- 2018 Cardiovascular and Metabolic Heterogeneity of Obesity. Clinical Challenges and Implications For ManagementDocument25 pages2018 Cardiovascular and Metabolic Heterogeneity of Obesity. Clinical Challenges and Implications For ManagementFrancisco Antonó Castro WeithNo ratings yet
- 2018 Evaluation of Spine MRIs in Athletes Participating in The Rio de Janeiro 2016 Summer Olympic GamesDocument8 pages2018 Evaluation of Spine MRIs in Athletes Participating in The Rio de Janeiro 2016 Summer Olympic GamesFrancisco Antonó Castro WeithNo ratings yet
- 2018 Match Running Performance in Professional Soccer Players Effect of Match Status and Goal DifferenceDocument3 pages2018 Match Running Performance in Professional Soccer Players Effect of Match Status and Goal DifferenceFrancisco Antonó Castro WeithNo ratings yet
- 2018 Muscle Strength Is A Poor Screening Test For Predicting Lower Extremity Injuries in Professional Male Soccer Players. A 2-Year Prospective Cohort StudyDocument11 pages2018 Muscle Strength Is A Poor Screening Test For Predicting Lower Extremity Injuries in Professional Male Soccer Players. A 2-Year Prospective Cohort StudyFrancisco Antonó Castro WeithNo ratings yet
- 2012 Recovery After High-Intensity Intermittent Exercise in Elite Soccer Players Using VEINOPLUS Sport Technology For Blood-Flow StimulationDocument9 pages2012 Recovery After High-Intensity Intermittent Exercise in Elite Soccer Players Using VEINOPLUS Sport Technology For Blood-Flow StimulationFrancisco Antonó Castro WeithNo ratings yet
- 2003 Hindlimb Unloading Alters Ligament HealingDocument11 pages2003 Hindlimb Unloading Alters Ligament HealingFrancisco Antonó Castro WeithNo ratings yet
- 2017 Genes To Predict VO2max Trainability A Systematic ReviewDocument30 pages2017 Genes To Predict VO2max Trainability A Systematic ReviewFrancisco Antonó Castro WeithNo ratings yet
- 2016 Genes and Athletic Performance An UpdateDocument15 pages2016 Genes and Athletic Performance An UpdateFrancisco Antonó Castro WeithNo ratings yet
- 2014 Kinematic Analysis of The Instep Kick in Youth Soccer PlayersDocument10 pages2014 Kinematic Analysis of The Instep Kick in Youth Soccer PlayersFrancisco Antonó Castro WeithNo ratings yet
- 1996 Effects of Moderate-Intensity Endurance and High-Intensity Intermittent Training On Anaerobic Capacity and VO (2max)Document12 pages1996 Effects of Moderate-Intensity Endurance and High-Intensity Intermittent Training On Anaerobic Capacity and VO (2max)Francisco Antonó Castro WeithNo ratings yet
- 2016 Effects of Low Versus High-Load Resistance Training On Muscle Strength and Hypertrophy in Well-Trained MenDocument32 pages2016 Effects of Low Versus High-Load Resistance Training On Muscle Strength and Hypertrophy in Well-Trained MenFrancisco Antonó Castro WeithNo ratings yet
- Human Movement Science: Full Length ArticleDocument10 pagesHuman Movement Science: Full Length ArticleFrancisco Antonó Castro WeithNo ratings yet
- 2016 Effects of Mental Imagery On Muscular Strength in Healthy and Patient Participant A Systematic ReviewDocument17 pages2016 Effects of Mental Imagery On Muscular Strength in Healthy and Patient Participant A Systematic ReviewFrancisco Antonó Castro WeithNo ratings yet
- 2016 Visceral Adiposity and Metabolic Syndrome After Very High-Fat and Low-Fat Isocaloric Diets A Randomized Controlled TrialDocument15 pages2016 Visceral Adiposity and Metabolic Syndrome After Very High-Fat and Low-Fat Isocaloric Diets A Randomized Controlled TrialFrancisco Antonó Castro WeithNo ratings yet
- 2017 Association of Weekend Warrior and Other Leisure Time Physical Activity Patterns With Risks For All-Cause, Cardiovascular Disease, and Cancer MortalityDocument8 pages2017 Association of Weekend Warrior and Other Leisure Time Physical Activity Patterns With Risks For All-Cause, Cardiovascular Disease, and Cancer MortalityFrancisco Antonó Castro WeithNo ratings yet
- 2017 Collateral Fattening When A Deficit in Lean Body Mass Drives OvereatingDocument3 pages2017 Collateral Fattening When A Deficit in Lean Body Mass Drives OvereatingFrancisco Antonó Castro WeithNo ratings yet
- 2016 Enhanced Muscle Insulin Sensitivity After Contraction-Exercise Is Mediated by AMPKDocument37 pages2016 Enhanced Muscle Insulin Sensitivity After Contraction-Exercise Is Mediated by AMPKFrancisco Antonó Castro WeithNo ratings yet
- 2016 Inverse Relationship Between Sleep Duration and Cardio-Ankle Vascular Index in ChildrenDocument8 pages2016 Inverse Relationship Between Sleep Duration and Cardio-Ankle Vascular Index in ChildrenFrancisco Antonó Castro WeithNo ratings yet
- 2009 Muscle Fatigue in Males and Females During Multiple-Sprint ExerciseDocument22 pages2009 Muscle Fatigue in Males and Females During Multiple-Sprint ExerciseFrancisco Antonó Castro WeithNo ratings yet
- 2016 Developmental Timing and Critical Windows For The Treatment of Psychiatric DisordersDocument10 pages2016 Developmental Timing and Critical Windows For The Treatment of Psychiatric DisordersFrancisco Antonó Castro WeithNo ratings yet
- Nina Fay Calhoun Award - Intl RelationsDocument5 pagesNina Fay Calhoun Award - Intl RelationsAltrusa International of Montrose CONo ratings yet
- The Good-Enough Sex Model For Couple Sexual SatisfactionDocument13 pagesThe Good-Enough Sex Model For Couple Sexual SatisfactionwernikNo ratings yet
- Grade 8 - P.E PPT (Week 1)Document30 pagesGrade 8 - P.E PPT (Week 1)Dave Sedigo100% (1)
- Final Sheet MotalityDocument69 pagesFinal Sheet MotalityAshima GabgotraNo ratings yet
- Mental Health Awareness SpeechDocument2 pagesMental Health Awareness SpeechThea Angela Longino95% (20)
- 2022 HRM Systems Diagnostic ChecklistsDocument5 pages2022 HRM Systems Diagnostic ChecklistsSacred EsportsNo ratings yet
- Part-IDocument507 pagesPart-INaan SivananthamNo ratings yet
- The Effect of Narcotic DrugDocument4 pagesThe Effect of Narcotic DrugFasra ChiongNo ratings yet
- Nurse - Resignation LetterDocument1 pageNurse - Resignation LetterphoenixdashNo ratings yet
- Receiving and Storage PDFDocument12 pagesReceiving and Storage PDFshyamkattiNo ratings yet
- Umbar - Performance Task 7Document3 pagesUmbar - Performance Task 7Bella CiaoNo ratings yet
- Job Safety Analysis Worksheet: JSA JSA Participants PPE Required Tools And/or EquipmentDocument5 pagesJob Safety Analysis Worksheet: JSA JSA Participants PPE Required Tools And/or EquipmentVigieNo ratings yet
- AishwaryaDocument52 pagesAishwaryamohitNo ratings yet
- FA Form No2-Visa Application FormDocument1 pageFA Form No2-Visa Application FormacademydonutNo ratings yet
- AICTE CorporateBestPracticesDocument13 pagesAICTE CorporateBestPracticesramar MNo ratings yet
- Wellness at SeaDocument9 pagesWellness at SeaRam Niwas ChauhanNo ratings yet
- A Solution-Focused Approach To Rational-Emotive Behavior Therapy - Toward A Theoretical IntegrationDocument22 pagesA Solution-Focused Approach To Rational-Emotive Behavior Therapy - Toward A Theoretical Integrationsolutions4familyNo ratings yet
- Gastritis - Etiology and Diagnosis - UpToDateDocument10 pagesGastritis - Etiology and Diagnosis - UpToDateLizbeth Navarrete SierraNo ratings yet
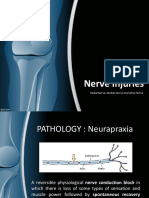
- Nerve Injuries: Radial Nerve, Median Nerve and Ulnar NerveDocument26 pagesNerve Injuries: Radial Nerve, Median Nerve and Ulnar NerveaisiyaahNo ratings yet
- Effective Leadership Towards The Star Rating Evaluation of Malaysian Seni Gayung Fatani Malaysia Organization PSGFMDocument10 pagesEffective Leadership Towards The Star Rating Evaluation of Malaysian Seni Gayung Fatani Malaysia Organization PSGFMabishekj274No ratings yet
- Tumors of Head and Neck RegionDocument94 pagesTumors of Head and Neck Regionpoornima vNo ratings yet
- Philippines AFHS - Standards and Implementation GuideDocument37 pagesPhilippines AFHS - Standards and Implementation GuideShardin Labawan-Juen,RNNo ratings yet
- EDC Annual ReportDocument433 pagesEDC Annual ReportAngela CanaresNo ratings yet
- World AIDS Day - December 1, 2019 Status of HIV Case-Based Surveillance Implementation - 39 U.S. PEPFAR-Supported Countries, May-July 2019Document16 pagesWorld AIDS Day - December 1, 2019 Status of HIV Case-Based Surveillance Implementation - 39 U.S. PEPFAR-Supported Countries, May-July 2019worksheetbookNo ratings yet
- Abc Ven 2020Document81 pagesAbc Ven 2020CorneLia JacintaNo ratings yet
- Lesson 4Document4 pagesLesson 4api-316910625No ratings yet
- YDA Nepal - MembersDocument13 pagesYDA Nepal - MembersSulochan LohaniNo ratings yet
- Monitoring The Performance of DamDocument137 pagesMonitoring The Performance of Damputih_138242459100% (2)
- Galay1 1 1 1Document2 pagesGalay1 1 1 1Glynne AlmadenNo ratings yet
- SUPW SrimukhDocument15 pagesSUPW SrimukhsrimukhsaiNo ratings yet