You might also like
- Contributors from Heriot-Watt University, Ryerson University, Scottish Water and moreDocument1 pageContributors from Heriot-Watt University, Ryerson University, Scottish Water and moreBudak Baru BelajarNo ratings yet
- Salasilah Raja Reman5Document1 pageSalasilah Raja Reman5Budak Baru BelajarNo ratings yet
- Webinar scriptsDocument2 pagesWebinar scriptsNurul FadilaNo ratings yet
- Chartered Scientist (CSci)Document1 pageChartered Scientist (CSci)Budak Baru BelajarNo ratings yet
- ARTIFICIAL DOUBLE PNEUMOTHORAX CASE STUDYDocument2 pagesARTIFICIAL DOUBLE PNEUMOTHORAX CASE STUDYBudak Baru BelajarNo ratings yet
- Waterborne Pathogens: Detection Methods and ApplicationsDocument1 pageWaterborne Pathogens: Detection Methods and ApplicationsBudak Baru BelajarNo ratings yet
- Comparison of Most Probable Number-PCR and Most Probable Number-Foci Detection Method For Quantifying Infectious Cryptosporidium Parvum Oocysts in Environmental SamplesDocument10 pagesComparison of Most Probable Number-PCR and Most Probable Number-Foci Detection Method For Quantifying Infectious Cryptosporidium Parvum Oocysts in Environmental SamplesBudak Baru BelajarNo ratings yet
- Comparison of Efficiency of Various DNA Extraction Methods From Cysts of Giardia Intestinalis Measured by PCR and TaqMan Real Time PCRDocument8 pagesComparison of Efficiency of Various DNA Extraction Methods From Cysts of Giardia Intestinalis Measured by PCR and TaqMan Real Time PCRBudak Baru BelajarNo ratings yet
- Sekolah Kurang MuridDocument11 pagesSekolah Kurang MuridBudak Baru BelajarNo ratings yet
- AcknowledgmentDocument1 pageAcknowledgmentBudak Baru BelajarNo ratings yet
- Handbook For Cell Culture Basics (Gibco)Document62 pagesHandbook For Cell Culture Basics (Gibco)Cesar PuentesNo ratings yet
- IndexDocument13 pagesIndexBudak Baru BelajarNo ratings yet
- Recent Advances in The Developmental Biology and Life Cycle of CryptosporidiumDocument13 pagesRecent Advances in The Developmental Biology and Life Cycle of CryptosporidiumBudak Baru BelajarNo ratings yet
- A Novel Photoinduced Electron Transfer (PET) Primer Technique For Rapid Real-Time PCR Detection of Cryptosporidium Spp.Document7 pagesA Novel Photoinduced Electron Transfer (PET) Primer Technique For Rapid Real-Time PCR Detection of Cryptosporidium Spp.Budak Baru BelajarNo ratings yet
- Cell Culture Basics HandbookDocument120 pagesCell Culture Basics HandbookbrunafariassNo ratings yet
- Comprehensive Guide to Unit Trust FundamentalsDocument132 pagesComprehensive Guide to Unit Trust FundamentalsMohd Hatif KamailNo ratings yet
- Cute Exam Paper Set 1Document20 pagesCute Exam Paper Set 1Budak Baru Belajar0% (1)
- 5 Reasons Cancer Researchers Adopt 3D Cell Culture White PaperDocument19 pages5 Reasons Cancer Researchers Adopt 3D Cell Culture White PaperBudak Baru BelajarNo ratings yet
- Cute Exam Paper Set 5Document12 pagesCute Exam Paper Set 5Budak Baru BelajarNo ratings yet
- Cute Exam Paper Set 4Document12 pagesCute Exam Paper Set 4Budak Baru BelajarNo ratings yet
- Introduction to Investment-Linked InsuranceDocument197 pagesIntroduction to Investment-Linked InsuranceSitisaiful0% (1)
- Karkun 9Document1 pageKarkun 9Budak Baru BelajarNo ratings yet
- Set 2 ENGLISH Question SolvedDocument19 pagesSet 2 ENGLISH Question SolvedBudak Baru BelajarNo ratings yet
- Cute Exam Paper Set 1Document20 pagesCute Exam Paper Set 1Budak Baru Belajar0% (1)
- 2013/14 Academic Calendar Year 1 Mbbs 220 Programme, Faculty of Medicine UitmDocument1 page2013/14 Academic Calendar Year 1 Mbbs 220 Programme, Faculty of Medicine UitmBudak Baru BelajarNo ratings yet
- Karkun 8Document1 pageKarkun 8Budak Baru BelajarNo ratings yet
- Cute Exam Paper Set 3Document18 pagesCute Exam Paper Set 3Budak Baru Belajar100% (1)
- Kitab Turath1Document11 pagesKitab Turath1Budak Baru Belajar100% (1)
- Medicinal OrchidsDocument14 pagesMedicinal OrchidsBudak Baru BelajarNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (587)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2219)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (119)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Administering Vaccines - Dose, Route, Site and Needle SizeDocument1 pageAdministering Vaccines - Dose, Route, Site and Needle SizeMark Jayson PecasalesNo ratings yet
- State of Illinois COVID-19 Update Nov. 6Document9 pagesState of Illinois COVID-19 Update Nov. 6CrainsChicagoBusinessNo ratings yet
- Case Study 1-LatumboDocument2 pagesCase Study 1-LatumboPao LatumboNo ratings yet
- Intestinal Nematodes Life CycleDocument7 pagesIntestinal Nematodes Life CyclealianaNo ratings yet
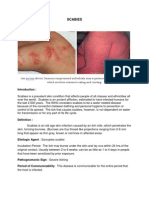
- ScabiesDocument8 pagesScabiesJessica Dumayas Dela RosaNo ratings yet
- Problem TreeDocument1 pageProblem Treeyel5buscatoNo ratings yet
- Anderson 2003 - Natural Histories of Infectious Disease - Ecological Vision in Twentieth-Century Biomedical ScienceDocument24 pagesAnderson 2003 - Natural Histories of Infectious Disease - Ecological Vision in Twentieth-Century Biomedical ScienceJerry ZeeNo ratings yet
- Malaria (Who)Document318 pagesMalaria (Who)RettyNo ratings yet
- DHFDocument40 pagesDHFPaulNo ratings yet
- Pestel Swot On Dairy IndDocument13 pagesPestel Swot On Dairy Indrudraksha sharmaNo ratings yet
- Grade3 Sense OrgansDocument28 pagesGrade3 Sense OrgansjindeylNo ratings yet
- Buildings and Structures: Padua EsteDocument2 pagesBuildings and Structures: Padua EsteKevin LavinaNo ratings yet
- Mosquito Bite PreventionDocument2 pagesMosquito Bite PreventionWSETNo ratings yet
- CAP76OCTDocument240 pagesCAP76OCTotorreslNo ratings yet
- 19th Century Safety Coffins Prevent Premature BurialsDocument5 pages19th Century Safety Coffins Prevent Premature BurialsBBDLNo ratings yet
- Ebola and MarburgDocument23 pagesEbola and Marburgtummalapalli venkateswara rao100% (1)
- Vpe 321 ZooDocument23 pagesVpe 321 ZooransinghNo ratings yet
- Critical Limb IschemiaDocument29 pagesCritical Limb IschemiadockunNo ratings yet
- Global DemographyDocument29 pagesGlobal DemographyLary Lou Ventura100% (4)
- Chapter15 StreptococciDocument66 pagesChapter15 StreptococciNursheda Abangon AzisNo ratings yet
- LetterDocument6 pagesLetteramanda mustikaNo ratings yet
- đề thi thử 2021 hàm rồngDocument8 pagesđề thi thử 2021 hàm rồngThu LeNo ratings yet
- ParasitologyDocument18 pagesParasitologyLudwig359100% (2)
- Diversity of MICROBIAL WORLDDocument47 pagesDiversity of MICROBIAL WORLDAdya Arradikha100% (1)
- Tippu Sultan Cover Story in AM - July 2012 FullDocument44 pagesTippu Sultan Cover Story in AM - July 2012 FullHarikumar NairNo ratings yet
- Kyoto Kagaku - Eye Examination Simulator PDFDocument2 pagesKyoto Kagaku - Eye Examination Simulator PDFJuan Carlos SuarezNo ratings yet
- Cholera Outbreak Guidelines: Preparedness, Prevention and ControlDocument105 pagesCholera Outbreak Guidelines: Preparedness, Prevention and ControlOxfamNo ratings yet
- Curriculum VitaeDocument3 pagesCurriculum VitaeLosta NataNo ratings yet
- Health Benefits of Figs or AnjeerDocument5 pagesHealth Benefits of Figs or AnjeerTaoshobuddhaNo ratings yet
- Quaratine ArchitectureDocument22 pagesQuaratine ArchitectureMarwa HassounNo ratings yet