You might also like
- 1 - 4922821657032130783 - CopiarDocument15 pages1 - 4922821657032130783 - CopiarLudmilaMerlinNo ratings yet
- Natural Innate and Adaptive Immunity To Cancer: FurtherDocument39 pagesNatural Innate and Adaptive Immunity To Cancer: FurtherSajjad Hossain ShuvoNo ratings yet
- Antimicrobial Activity of Nanoparticles: Applications in Wound Healing and Infection TreatmentFrom EverandAntimicrobial Activity of Nanoparticles: Applications in Wound Healing and Infection TreatmentGregory GuisbiersNo ratings yet
- Kaufmann Immunology of Infection 2010Document541 pagesKaufmann Immunology of Infection 2010Larisa StanNo ratings yet
- Behavioral and Department ModelsFrom EverandBehavioral and Department ModelsBert ZuckermanNo ratings yet
- Tested Studies For Laboratory Teaching. Peifer. 1989Document174 pagesTested Studies For Laboratory Teaching. Peifer. 1989NidzaNo ratings yet
- Lab FaqsDocument384 pagesLab FaqsPriyanka SinghNo ratings yet
- Manual PDFDocument53 pagesManual PDFPrashanth N SuravajhalaNo ratings yet
- Enzyme Inhibition in Drug Discovery and Development: The Good and the BadFrom EverandEnzyme Inhibition in Drug Discovery and Development: The Good and the BadChuang LuNo ratings yet
- Mangel - The Theoretical Biologist's Toolbox - Quantitative Methods For Ecology and Evolutionary Biology (Cambridge, 2006) PDFDocument391 pagesMangel - The Theoretical Biologist's Toolbox - Quantitative Methods For Ecology and Evolutionary Biology (Cambridge, 2006) PDFhightdante100% (2)
- Biosimilars of Monoclonal Antibodies: A Practical Guide to Manufacturing, Preclinical, and Clinical DevelopmentFrom EverandBiosimilars of Monoclonal Antibodies: A Practical Guide to Manufacturing, Preclinical, and Clinical DevelopmentNo ratings yet
- Whitepaper Benefits DNA Sequencing Microbial Identification Pharma Analytics June 2022Document5 pagesWhitepaper Benefits DNA Sequencing Microbial Identification Pharma Analytics June 2022JohnNo ratings yet
- Recent Advancements in Microbial DiversityFrom EverandRecent Advancements in Microbial DiversitySurajit de MandalNo ratings yet
- Biotechnology in The Diagnosis of Infectious Diseases and Vaccine DevelopmentDocument25 pagesBiotechnology in The Diagnosis of Infectious Diseases and Vaccine DevelopmentRahila IqbalNo ratings yet
- Searching For Hypothetical ProteinsDocument38 pagesSearching For Hypothetical Proteinssumera120488No ratings yet
- In Vitro Methods For Detecting Cytotoxicity: UNIT 2.6Document27 pagesIn Vitro Methods For Detecting Cytotoxicity: UNIT 2.6hiền nguyễnNo ratings yet
- Heterotrophic Planktonic Microbes: Virus, Bacteria, Archaea, and ProtozoaDocument34 pagesHeterotrophic Planktonic Microbes: Virus, Bacteria, Archaea, and ProtozoaCharity MbiziNo ratings yet
- Strategies For Culture of Unculturable BacteriaDocument7 pagesStrategies For Culture of Unculturable BacteriaEd M. K-zhan NözNo ratings yet
- Antony Van Leeuw en 00 Do BeDocument516 pagesAntony Van Leeuw en 00 Do BeRonNo ratings yet
- Salmonella - Methods and Protocols.-springer-Verlag New York (2020)Document215 pagesSalmonella - Methods and Protocols.-springer-Verlag New York (2020)Yudith ParamitadeviNo ratings yet
- FungusDocument31 pagesFungushadiya rehmanNo ratings yet
- GTI Manual Final 25juneDocument135 pagesGTI Manual Final 25juneoscarnicolasNo ratings yet
- Science Magazine 5857 2007-12-14Document97 pagesScience Magazine 5857 2007-12-14sumersoft1No ratings yet
- Goriely 2017Document652 pagesGoriely 2017Тимофей НестеровNo ratings yet
- Manual On Antimicrobial Susceptibility Testing Dr. M.K. LalithaDocument47 pagesManual On Antimicrobial Susceptibility Testing Dr. M.K. LalithaAntoniuz Liem100% (2)
- Science Magazine 5693 2004-10-01Document107 pagesScience Magazine 5693 2004-10-01WillimSmith100% (1)
- Moving Environmental DNA Methods From Concept To Practice For Monitoring Aquatic MacroorganismsDocument3 pagesMoving Environmental DNA Methods From Concept To Practice For Monitoring Aquatic MacroorganismsFrancisco PrietoNo ratings yet
- Resazurin Cell Viability Assay - Orzel (Full)Document5 pagesResazurin Cell Viability Assay - Orzel (Full)ShawnNo ratings yet
- Candida Glabrata, Candida Parapsilosis Andcandida Tropicalis PDFDocument18 pagesCandida Glabrata, Candida Parapsilosis Andcandida Tropicalis PDFNovena DpNo ratings yet
- Enterococci AgarDocument2 pagesEnterococci Agartevy101No ratings yet
- M Endo Broth MF: Intended Use FormulaDocument2 pagesM Endo Broth MF: Intended Use Formulasami algradeeNo ratings yet
- Laboratory Manual: 1 Lmbty360Document50 pagesLaboratory Manual: 1 Lmbty360Rise from the ashes100% (1)
- A Microbial Loop-Ocn Bio 1Document6 pagesA Microbial Loop-Ocn Bio 1Fagner RicardoNo ratings yet
- SOP For Antimicrobial Effectiveness TestingDocument4 pagesSOP For Antimicrobial Effectiveness TestingGencay ErginNo ratings yet
- 2019 Book PolymyxinAntibioticsFromLaboraDocument364 pages2019 Book PolymyxinAntibioticsFromLaboraDaniel JiménezNo ratings yet
- HSC Biotech OptionDocument3 pagesHSC Biotech OptionthalwiNo ratings yet
- Applications of Aptamers As SensorsDocument26 pagesApplications of Aptamers As SensorsAndrey ZdanovichNo ratings yet
- Laboratory ManualDocument32 pagesLaboratory ManuallkjwrotuNo ratings yet
- Escherichia Coli (All Strains) Quantification Of: Uida (Glucuronidase)Document10 pagesEscherichia Coli (All Strains) Quantification Of: Uida (Glucuronidase)khoa151290No ratings yet
- ATCC Strains and Papers Quality Control Solutions For Food TestingDocument12 pagesATCC Strains and Papers Quality Control Solutions For Food Testingernesto arriazaNo ratings yet
- The EZ-Product Family: Make Your Microbiology Workflow EasyDocument8 pagesThe EZ-Product Family: Make Your Microbiology Workflow Easyaisyah annisa rahma hidayahNo ratings yet
- Crispr-Cas Systems For Editing, Regulating and Targeting GenomesDocument9 pagesCrispr-Cas Systems For Editing, Regulating and Targeting GenomesToman Chi To LamNo ratings yet
- Book - Mathematics - Theoretical - Biology PDFDocument130 pagesBook - Mathematics - Theoretical - Biology PDFIsrael Castillo100% (1)
- Probiotics-From Metchnikoff To BioactivesDocument15 pagesProbiotics-From Metchnikoff To Bioactivesmilu1312No ratings yet
- Pick The Right One at A Glance: The New Color-Coded MC-Media PadsDocument6 pagesPick The Right One at A Glance: The New Color-Coded MC-Media PadsAji HutamaNo ratings yet
- Molds in Museum Environments: Biodeterioration of Art Photographs and Wooden SculpturesDocument8 pagesMolds in Museum Environments: Biodeterioration of Art Photographs and Wooden SculpturesOPTICALMIMOOFDMNo ratings yet
- Universal GEN III Protocol 07172013Document6 pagesUniversal GEN III Protocol 07172013Ashraf ElmougiNo ratings yet
- Biomimetics - A Molecular PerspectiveDocument260 pagesBiomimetics - A Molecular PerspectivePeterAttilaMarcoNo ratings yet
- Cell Proliferation Kit I (MTT) : Cat. No. 11 465 007 001Document4 pagesCell Proliferation Kit I (MTT) : Cat. No. 11 465 007 001AyuAnatrieraNo ratings yet
- MC Media Pad BrochureDocument6 pagesMC Media Pad Brochurerosalia destikaNo ratings yet
- (Advances in Experimental Medicine and Biology 897) Gianfranco Donelli (Eds.) - Advances in Microbiology, Infectious Diseases and Public Health - Volume 1-Springer International Publishing (2016)Document128 pages(Advances in Experimental Medicine and Biology 897) Gianfranco Donelli (Eds.) - Advances in Microbiology, Infectious Diseases and Public Health - Volume 1-Springer International Publishing (2016)niaNo ratings yet
- Protoplast Culture and Somatic HybridizationDocument14 pagesProtoplast Culture and Somatic HybridizationRollor CosterNo ratings yet
- Microbial Diversity Manual PDFDocument83 pagesMicrobial Diversity Manual PDFCesar AlegreNo ratings yet
- 3M Petrifilm Yeast MoldsDocument8 pages3M Petrifilm Yeast MoldsMelvin BendicioNo ratings yet
- Compact - Dry - Catalog 2022Document11 pagesCompact - Dry - Catalog 2022Gabriela CalderonNo ratings yet
- Case Report: Diastema Closure in Anterior Teeth Using A Posterior MatrixDocument7 pagesCase Report: Diastema Closure in Anterior Teeth Using A Posterior MatrixJing XueNo ratings yet
- 10.2319 080619-516EPubDocument8 pages10.2319 080619-516EPubJing XueNo ratings yet
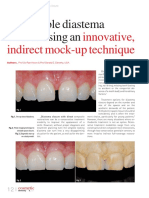
- Innovative, Indirect Mock-Up Technique: Predictable Diastema Closure Using AnDocument4 pagesInnovative, Indirect Mock-Up Technique: Predictable Diastema Closure Using AnJing XueNo ratings yet
- Intraorifice Sealing Ability of Different Materials in Endodontically Treated TeethDocument5 pagesIntraorifice Sealing Ability of Different Materials in Endodontically Treated TeethJing XueNo ratings yet
- Incidence of Pain Associated With Clinical Factors During and After Root Canal Therapy. Part 1. Interappointment PainDocument4 pagesIncidence of Pain Associated With Clinical Factors During and After Root Canal Therapy. Part 1. Interappointment PainJing XueNo ratings yet
- Closure of Diastema and Gingival Recontouring Using Direct Adhesive Restorations: A Case ReportDocument12 pagesClosure of Diastema and Gingival Recontouring Using Direct Adhesive Restorations: A Case ReportJing XueNo ratings yet
- 0003-3219 (1977) 047 0265 Tmidai 2 0 Co 2Document7 pages0003-3219 (1977) 047 0265 Tmidai 2 0 Co 2Jing XueNo ratings yet
- Avulsions and Intrusions: The Controversial Displacement InjuriesDocument7 pagesAvulsions and Intrusions: The Controversial Displacement InjuriesJing XueNo ratings yet
- Survival of Restored Endodontically Treated Teeth in Relation To Periodontal StatusDocument4 pagesSurvival of Restored Endodontically Treated Teeth in Relation To Periodontal StatusJing XueNo ratings yet
- Project Deliverable: Safety and Efficacy of A New and Emerging Dental X-Ray ModalityDocument16 pagesProject Deliverable: Safety and Efficacy of A New and Emerging Dental X-Ray ModalityJing XueNo ratings yet
- Walton SinusTractTracingEffectivenessDocument1 pageWalton SinusTractTracingEffectivenessJing XueNo ratings yet
- Outcome of Root Canal Treatment in Dogs Determined by Periapical Radiography and Cone-Beam Computed Tomography ScansDocument4 pagesOutcome of Root Canal Treatment in Dogs Determined by Periapical Radiography and Cone-Beam Computed Tomography ScansJing XueNo ratings yet
- BRONJinDentalImplantPatientsGossAustralia PDFDocument7 pagesBRONJinDentalImplantPatientsGossAustralia PDFJing XueNo ratings yet
- Why Clinicians Are Natural Bayesians: Interpreting Diagnostic Test Results From The Bayesian PerspectiveDocument4 pagesWhy Clinicians Are Natural Bayesians: Interpreting Diagnostic Test Results From The Bayesian PerspectiveJing XueNo ratings yet
- Dental Vertical Root Fractures: Value of CT in Detection: Head and Neck ImagingDocument5 pagesDental Vertical Root Fractures: Value of CT in Detection: Head and Neck ImagingJing XueNo ratings yet
- The AAE's Endodontic Case Difficulty Assessment Form Offers AnswersDocument2 pagesThe AAE's Endodontic Case Difficulty Assessment Form Offers AnswersJing XueNo ratings yet
- Abilityofcone beamcomputerizedtomographytodetectVRFcasereport2011Document5 pagesAbilityofcone beamcomputerizedtomographytodetectVRFcasereport2011Jing XueNo ratings yet
- Wide Diameter Implants-Degidi Piattelli LezziDocument7 pagesWide Diameter Implants-Degidi Piattelli LezziJing XueNo ratings yet
- AAOMR Cone Beam PaperDocument2 pagesAAOMR Cone Beam PaperJing XueNo ratings yet
- User S Manual AURORA 1.2K - 2.2KDocument288 pagesUser S Manual AURORA 1.2K - 2.2KEprom ServisNo ratings yet
- Digital MetersDocument47 pagesDigital MetersherovhungNo ratings yet
- C C C C: "P P P P PDocument25 pagesC C C C: "P P P P PShalu Dua KatyalNo ratings yet
- User Manual For Speed Control of BLDC Motor Using DspicDocument12 pagesUser Manual For Speed Control of BLDC Motor Using DspicTrung TrựcNo ratings yet
- 1.technical Specifications (Piling)Document15 pages1.technical Specifications (Piling)Kunal Panchal100% (2)
- ISBN Safe Work Method Statements 2022 03Document8 pagesISBN Safe Work Method Statements 2022 03Tamo Kim ChowNo ratings yet
- Hdfs Default XML ParametersDocument14 pagesHdfs Default XML ParametersVinod BihalNo ratings yet
- Developing Paragraphs: Test Section - Writing Task 2Document11 pagesDeveloping Paragraphs: Test Section - Writing Task 2Cengizhan AkdağNo ratings yet
- Role of Personal Finance Towards Managing of Money - DraftaDocument35 pagesRole of Personal Finance Towards Managing of Money - DraftaAndrea Denise Lion100% (1)
- 7400 IC SeriesDocument16 pages7400 IC SeriesRaj ZalariaNo ratings yet
- Fss Presentation Slide GoDocument13 pagesFss Presentation Slide GoReinoso GreiskaNo ratings yet
- DPSD ProjectDocument30 pagesDPSD ProjectSri NidhiNo ratings yet
- Chapter3 Elasticity and ForecastingDocument25 pagesChapter3 Elasticity and ForecastingGee JoeNo ratings yet
- Ozone Therapy - A Clinical Review A. M. Elvis and J. S. EktaDocument5 pagesOzone Therapy - A Clinical Review A. M. Elvis and J. S. Ektatahuti696No ratings yet
- 11-03 TB Value Chains and BPs - WolfDocument3 pages11-03 TB Value Chains and BPs - WolfPrakash PandeyNo ratings yet
- Excon2019 ShowPreview02122019 PDFDocument492 pagesExcon2019 ShowPreview02122019 PDFSanjay KherNo ratings yet
- Crown WF-3000 1.2Document5 pagesCrown WF-3000 1.2Qirat KhanNo ratings yet
- Solved Simplex Problems PDFDocument5 pagesSolved Simplex Problems PDFTejasa MishraNo ratings yet
- Ricoh IM C2000 IM C2500: Full Colour Multi Function PrinterDocument4 pagesRicoh IM C2000 IM C2500: Full Colour Multi Function PrinterKothapalli ChiranjeeviNo ratings yet
- Elements of ArtDocument1 pageElements of Artsamson8cindy8louNo ratings yet
- Azimuth Steueung - EngDocument13 pagesAzimuth Steueung - EnglacothNo ratings yet
- Retailing in IndiaDocument11 pagesRetailing in IndiaVinod MalkarNo ratings yet
- Week 7 Sex Limited InfluencedDocument19 pagesWeek 7 Sex Limited InfluencedLorelyn VillamorNo ratings yet
- Agco Serie 800 PDFDocument24 pagesAgco Serie 800 PDFJohnny VargasNo ratings yet
- HirePro Video Proctored Online-Instruction Sheet - Bain IndiaDocument1 pageHirePro Video Proctored Online-Instruction Sheet - Bain Indiaapoorv sharmaNo ratings yet
- Assignment#10 Global Strategy and The Multinational CorporationDocument1 pageAssignment#10 Global Strategy and The Multinational CorporationAnjaneth A. VillegasNo ratings yet
- WebLMT HelpDocument12 pagesWebLMT HelpJoão LopesNo ratings yet
- Emea 119948060Document31 pagesEmea 119948060ASHUTOSH MISHRANo ratings yet
- TCL LD24D50 - Chassis MS09A-LA - (TKLE2413D) - Manual de Servicio PDFDocument41 pagesTCL LD24D50 - Chassis MS09A-LA - (TKLE2413D) - Manual de Servicio PDFFabian OrtuzarNo ratings yet
- Lesson 6 ComprogDocument25 pagesLesson 6 ComprogmarkvillaplazaNo ratings yet
- 10% Human: How Your Body's Microbes Hold the Key to Health and HappinessFrom Everand10% Human: How Your Body's Microbes Hold the Key to Health and HappinessRating: 4 out of 5 stars4/5 (33)
- The Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceFrom EverandThe Molecule of More: How a Single Chemical in Your Brain Drives Love, Sex, and Creativity--and Will Determine the Fate of the Human RaceRating: 4.5 out of 5 stars4.5/5 (517)
- Why We Die: The New Science of Aging and the Quest for ImmortalityFrom EverandWhy We Die: The New Science of Aging and the Quest for ImmortalityRating: 4 out of 5 stars4/5 (5)
- Tales from Both Sides of the Brain: A Life in NeuroscienceFrom EverandTales from Both Sides of the Brain: A Life in NeuroscienceRating: 3 out of 5 stars3/5 (18)
- Return of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseFrom EverandReturn of the God Hypothesis: Three Scientific Discoveries That Reveal the Mind Behind the UniverseRating: 4.5 out of 5 stars4.5/5 (52)
- When the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisFrom EverandWhen the Body Says No by Gabor Maté: Key Takeaways, Summary & AnalysisRating: 3.5 out of 5 stars3.5/5 (2)
- Undeniable: How Biology Confirms Our Intuition That Life Is DesignedFrom EverandUndeniable: How Biology Confirms Our Intuition That Life Is DesignedRating: 4 out of 5 stars4/5 (11)
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (393)
- The Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionFrom EverandThe Ancestor's Tale: A Pilgrimage to the Dawn of EvolutionRating: 4 out of 5 stars4/5 (812)
- All That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesFrom EverandAll That Remains: A Renowned Forensic Scientist on Death, Mortality, and Solving CrimesRating: 4.5 out of 5 stars4.5/5 (397)
- A Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsFrom EverandA Brief History of Intelligence: Evolution, AI, and the Five Breakthroughs That Made Our BrainsRating: 4.5 out of 5 stars4.5/5 (6)
- Good Without God: What a Billion Nonreligious People Do BelieveFrom EverandGood Without God: What a Billion Nonreligious People Do BelieveRating: 4 out of 5 stars4/5 (66)
- Seven and a Half Lessons About the BrainFrom EverandSeven and a Half Lessons About the BrainRating: 4 out of 5 stars4/5 (110)
- Buddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomFrom EverandBuddha's Brain: The Practical Neuroscience of Happiness, Love & WisdomRating: 4 out of 5 stars4/5 (216)
- Who's in Charge?: Free Will and the Science of the BrainFrom EverandWho's in Charge?: Free Will and the Science of the BrainRating: 4 out of 5 stars4/5 (65)
- The Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorFrom EverandThe Other Side of Normal: How Biology Is Providing the Clues to Unlock the Secrets of Normal and Abnormal BehaviorNo ratings yet
- The Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindFrom EverandThe Consciousness Instinct: Unraveling the Mystery of How the Brain Makes the MindRating: 4.5 out of 5 stars4.5/5 (93)
- Gut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)From EverandGut: The Inside Story of Our Body's Most Underrated Organ (Revised Edition)Rating: 4 out of 5 stars4/5 (411)
- The Lives of Bees: The Untold Story of the Honey Bee in the WildFrom EverandThe Lives of Bees: The Untold Story of the Honey Bee in the WildRating: 4.5 out of 5 stars4.5/5 (44)
- Darwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignFrom EverandDarwin's Doubt: The Explosive Origin of Animal Life and the Case for Intelligent DesignRating: 4.5 out of 5 stars4.5/5 (39)
- The Rise and Fall of the Dinosaurs: A New History of a Lost WorldFrom EverandThe Rise and Fall of the Dinosaurs: A New History of a Lost WorldRating: 4 out of 5 stars4/5 (597)
- Moral Tribes: Emotion, Reason, and the Gap Between Us and ThemFrom EverandMoral Tribes: Emotion, Reason, and the Gap Between Us and ThemRating: 4.5 out of 5 stars4.5/5 (115)
- Human: The Science Behind What Makes Your Brain UniqueFrom EverandHuman: The Science Behind What Makes Your Brain UniqueRating: 3.5 out of 5 stars3.5/5 (38)
- Fast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperFrom EverandFast Asleep: Improve Brain Function, Lose Weight, Boost Your Mood, Reduce Stress, and Become a Better SleeperRating: 4.5 out of 5 stars4.5/5 (16)