You might also like
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (844)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5810)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (347)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Multiplication 5 Minute Frenzy (A)Document1 pageMultiplication 5 Minute Frenzy (A)NaomiKerryaThompsonNo ratings yet
- Biology PDFDocument1,447 pagesBiology PDFPrincess Ethel Atillo100% (2)
- Solution Manual For Nesters Microbiology A Human Perspective 10th Edition Denise Anderson Sarah Salm Eugene NesterDocument9 pagesSolution Manual For Nesters Microbiology A Human Perspective 10th Edition Denise Anderson Sarah Salm Eugene NesterChristianColemannsja100% (38)
- Digital EvidenceDocument22 pagesDigital EvidenceNaomiKerryaThompsonNo ratings yet
- (Learning Materials in Biosciences) Melanie Kappelmann-Fenzl - Next Generation Sequencing and Data Analysis-Springer (2021)Document225 pages(Learning Materials in Biosciences) Melanie Kappelmann-Fenzl - Next Generation Sequencing and Data Analysis-Springer (2021)gndfbj100% (2)
- Dukewriting Lesson3-AnswerDocument2 pagesDukewriting Lesson3-AnswerSalekin MishuNo ratings yet
- Standard Five Pupil Aces CSEC Math - Grade ADocument2 pagesStandard Five Pupil Aces CSEC Math - Grade ANaomiKerryaThompsonNo ratings yet
- UNIT 7 Number System and Bases: Introduction To SIMDocument2 pagesUNIT 7 Number System and Bases: Introduction To SIMNaomiKerryaThompsonNo ratings yet
- Multiplication 5 Minute Frenzy (B)Document1 pageMultiplication 5 Minute Frenzy (B)NaomiKerryaThompsonNo ratings yet
- Maths Review TestDocument5 pagesMaths Review TestNaomiKerryaThompsonNo ratings yet
- Maths Review TestDocument5 pagesMaths Review TestNaomiKerryaThompsonNo ratings yet
- Sea Guidelines PDFDocument28 pagesSea Guidelines PDFNaomiKerryaThompsonNo ratings yet
- Sea Guidelines PDFDocument28 pagesSea Guidelines PDFNaomiKerryaThompsonNo ratings yet
- Application Form PSTA PostsDocument4 pagesApplication Form PSTA PostsNaomiKerryaThompsonNo ratings yet
- Times Table 1-4Document1 pageTimes Table 1-4NaomiKerryaThompsonNo ratings yet
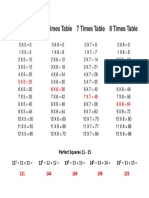
- Times Table 5-8Document1 pageTimes Table 5-8NaomiKerryaThompsonNo ratings yet
- Multiplication & Division With NegativesDocument2 pagesMultiplication & Division With NegativesNaomiKerryaThompsonNo ratings yet
- Maths Review TestDocument5 pagesMaths Review TestNaomiKerryaThompsonNo ratings yet
- Division Tables Table 1-4Document1 pageDivision Tables Table 1-4NaomiKerryaThompsonNo ratings yet
- Color Vision 3Document14 pagesColor Vision 3NaomiKerryaThompsonNo ratings yet
- Ni Hms 51627Document32 pagesNi Hms 51627NaomiKerryaThompsonNo ratings yet
- Nucleolus A Central Hub For Nuclear FunctionsDocument13 pagesNucleolus A Central Hub For Nuclear FunctionsJuan Andres Fernández MNo ratings yet
- Monalisa ManurungDocument13 pagesMonalisa ManurungPradinta BayuNo ratings yet
- What Are Introns and Exons - PDFDocument14 pagesWhat Are Introns and Exons - PDFSterlingNo ratings yet
- Protein Synthesis2Document56 pagesProtein Synthesis2api-248442486No ratings yet
- ZoologyDocument46 pagesZoologyRahul Kumar SharmaNo ratings yet
- Gene RegulationDocument37 pagesGene RegulationJonas T. HingcoNo ratings yet
- Science - Abo7257 SMDocument105 pagesScience - Abo7257 SM张议No ratings yet
- From Green Plants To Industrial EnzymesDocument5 pagesFrom Green Plants To Industrial Enzymesricky_connollyNo ratings yet
- HAYATI Journal of Biosciences: Fenny M. Dwivany, Rizkita Rahmi Esyanti, Aksarani 'Sa Pratiwi, Hera Fi ZaskiaDocument4 pagesHAYATI Journal of Biosciences: Fenny M. Dwivany, Rizkita Rahmi Esyanti, Aksarani 'Sa Pratiwi, Hera Fi ZaskiaCherylou CamusNo ratings yet
- Biotechnology QuizDocument3 pagesBiotechnology QuizDwi Septiani PutriNo ratings yet
- HEC Revised Biotech SyllabusDocument56 pagesHEC Revised Biotech SyllabuswaheedgulNo ratings yet
- CENTRAL DOGMA Translation Transcription Protein SynthesisDocument21 pagesCENTRAL DOGMA Translation Transcription Protein SynthesisCristie Ann GuiamNo ratings yet
- Syllabus For 5-Years Integrated M. Sc. Degree: UM-DAE Centre For Excellence in Basic SciencesDocument92 pagesSyllabus For 5-Years Integrated M. Sc. Degree: UM-DAE Centre For Excellence in Basic SciencesAubal ShaubalNo ratings yet
- 6.2 D Protein Synthesis: ContosoDocument13 pages6.2 D Protein Synthesis: Contosoruva gundaNo ratings yet
- Chapter 11 Gene ExpressionDocument30 pagesChapter 11 Gene ExpressionJonathan WyattNo ratings yet
- 12 Biology Notes Ch06 Molecular Basis of InheritanceDocument6 pages12 Biology Notes Ch06 Molecular Basis of InheritanceInderpal SinghNo ratings yet
- (PDF) Engineering Disease Resistant CattleDocument7 pages(PDF) Engineering Disease Resistant CattlecarlesNo ratings yet
- BioCentralDogmaWorksheet 1Document3 pagesBioCentralDogmaWorksheet 1jaelajeanine326No ratings yet
- Physrev 00036 2015Document34 pagesPhysrev 00036 2015Sinisa RisticNo ratings yet
- Various Expression SystemDocument15 pagesVarious Expression SystemVamsi K KuchibhotlaNo ratings yet
- Dual Luciferase Reporter Assay System Protocol PDFDocument26 pagesDual Luciferase Reporter Assay System Protocol PDFThanos86No ratings yet
- Bio 1002 B Mar FeedbackDocument13 pagesBio 1002 B Mar FeedbackLouise2880No ratings yet
- Mechanisms of Epigenetic Inheritance: Cyrus Martin and Yi ZhangDocument7 pagesMechanisms of Epigenetic Inheritance: Cyrus Martin and Yi Zhangbahatska2No ratings yet
- Signal HypothesisDocument3 pagesSignal Hypothesischandra mouliNo ratings yet
- Course Outline 2280A 2023 FinalDocument9 pagesCourse Outline 2280A 2023 Finalmya tararNo ratings yet
- Protein Synthesis Extra CreditDocument2 pagesProtein Synthesis Extra Creditcmillica1176No ratings yet