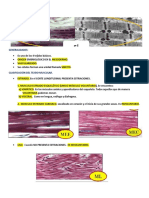
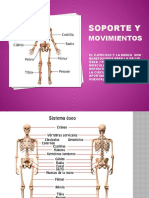
You might also like
- Clostridium BotulinumDocument2 pagesClostridium BotulinumchabelongaNo ratings yet
- ATUNDocument2 pagesATUNCat AsassinNo ratings yet
- Sistema Nerviosos Central y PeriféricoDocument11 pagesSistema Nerviosos Central y PeriféricoJoselyn LindoNo ratings yet
- Caso ClínicoDocument3 pagesCaso ClínicoIvonne VillegasNo ratings yet
- Esquema-Resumen Bloque MetabolismoDocument38 pagesEsquema-Resumen Bloque MetabolismolauraNo ratings yet
- Metabolismo Hidratos de CarbonoDocument25 pagesMetabolismo Hidratos de CarbonoSergio Sanchez LabarcaNo ratings yet
- Certificado BerkleeX OMPRD-100x - EdXDocument1 pageCertificado BerkleeX OMPRD-100x - EdXM.C. RidulfoNo ratings yet
- OrganogenesisDocument100 pagesOrganogenesisPedro RojasNo ratings yet
- Alimentación de TernerosDocument55 pagesAlimentación de TernerosJaime Sangay CotrinaNo ratings yet
- ORGANELOS de La Célula AnimalDocument15 pagesORGANELOS de La Célula AnimalmaylinNo ratings yet
- Ojo Anatomía e Histología, FinalDocument6 pagesOjo Anatomía e Histología, FinalJacob MendozaNo ratings yet
- Organelos de La Celula Procariota AnimalDocument64 pagesOrganelos de La Celula Procariota AnimalOmar OrdoñezNo ratings yet
- Anatomia Del CorazónDocument1 pageAnatomia Del CorazónNathalie DávilaNo ratings yet
- Genealogías y Tipos de HerenciaDocument20 pagesGenealogías y Tipos de HerenciaRicardo Álvarez BernalNo ratings yet
- Ciclo de KrebsDocument8 pagesCiclo de KrebsGustavo Adolfo MozNo ratings yet
- Articulaciones y tipos de movimientoDocument47 pagesArticulaciones y tipos de movimientobhanny2No ratings yet
- Tema 1 Citología e Histología VeterinariaDocument21 pagesTema 1 Citología e Histología VeterinariaCorreoNo ratings yet
- Leguminosas espontaneas de posible valor forrajero en ColombiaFrom EverandLeguminosas espontaneas de posible valor forrajero en ColombiaNo ratings yet
- Proteínas globulares vs fibrosasDocument3 pagesProteínas globulares vs fibrosasCamilo Cortina AlfaroNo ratings yet
- Metabolismo proteico en animalesDocument26 pagesMetabolismo proteico en animalesNatalia LopezNo ratings yet
- MINERALES Y VITAMINAS EN RUMIANTES Y NO RUMIANTESDocument28 pagesMINERALES Y VITAMINAS EN RUMIANTES Y NO RUMIANTESroberth paulNo ratings yet
- Tipos de EpitelioDocument7 pagesTipos de EpitelioYoscaryDeLastraLicettNo ratings yet
- Digestión de ProteinasDocument16 pagesDigestión de ProteinasLaryssa PedraNo ratings yet
- Conejo Hamster DDocument2 pagesConejo Hamster DAlmendra E. Henriquez VargasNo ratings yet
- Miología Del Cráneo EquinoDocument8 pagesMiología Del Cráneo Equinojohan_perez900% (1)
- Bioquimica TamponesDocument13 pagesBioquimica TamponesrmdlcNo ratings yet
- Epidermitis ExudativaDocument4 pagesEpidermitis ExudativaloboleunamNo ratings yet
- Produccion PorcinaDocument12 pagesProduccion PorcinaJorge Luis Ramirez BenitezNo ratings yet
- Sistemas de Produccion AnimalDocument24 pagesSistemas de Produccion AnimalAndrea Santamaria100% (1)
- Diagnóstico de gestación en bovinosDocument23 pagesDiagnóstico de gestación en bovinosDanilo Arenas100% (1)
- Parte - VIII Sistema EndocrinoDocument192 pagesParte - VIII Sistema Endocrinovaleria restrepoNo ratings yet
- Nom Del Expediente Clínico 004Document15 pagesNom Del Expediente Clínico 004LizethAlvidrez100% (1)
- Regulacion y Tolerancia Sistema InmuneDocument37 pagesRegulacion y Tolerancia Sistema InmuneAna María RodrigoNo ratings yet
- Primeros Pensadores ExistencialesDocument2 pagesPrimeros Pensadores ExistencialesOliva Guevara FredyNo ratings yet
- ProteinasDocument6 pagesProteinasRodrigo LopezNo ratings yet
- Laminitis EquinaDocument7 pagesLaminitis EquinaAlex Gabriel Soto PintoNo ratings yet
- Anatomía descriptiva de animalesDocument15 pagesAnatomía descriptiva de animalesCarlos Fernando Zambrano Triana50% (2)
- Biología de Los MicroorganismosDocument4 pagesBiología de Los MicroorganismosGiovanni MoralesNo ratings yet
- EukaryotaDocument8 pagesEukaryotaOscar Alejandro Escobedo Nevarez100% (1)
- Pietin Ovino y CaprinoDocument14 pagesPietin Ovino y Caprinoapuntesdelcev0% (1)
- Apunte Sistema Nervioso Autónomo (1) VeterinariaDocument10 pagesApunte Sistema Nervioso Autónomo (1) VeterinariaNicolas SaraccoNo ratings yet
- Deficiencia de La Vitamina C en CobayosDocument4 pagesDeficiencia de La Vitamina C en CobayosGuadalupe Rosario TarquiNo ratings yet
- Giardia LambliaDocument14 pagesGiardia LambliaJORDY100% (1)
- Displasia de codo en perros: anatomía y patologíaDocument13 pagesDisplasia de codo en perros: anatomía y patologíajuan mateusNo ratings yet
- CascoDocument14 pagesCascoAgus DíazNo ratings yet
- Glándulas y SecreciónDocument26 pagesGlándulas y SecreciónMariana VillarrealNo ratings yet
- Caso Clinico Veterinaria NeoplasiasDocument4 pagesCaso Clinico Veterinaria NeoplasiasSandie SaldivaarNo ratings yet
- Parto y nacimiento: evolución histórica y etapas del procesoDocument14 pagesParto y nacimiento: evolución histórica y etapas del procesoPamela CarlosNo ratings yet
- Alimentación Del Toro de LidiaDocument24 pagesAlimentación Del Toro de LidiaNestor E. ObispoNo ratings yet
- PLDocument115 pagesPLAnonymous LANCVGRE8No ratings yet
- Tecnica de Necropsia en Perros PDFDocument16 pagesTecnica de Necropsia en Perros PDFJare NolascoNo ratings yet
- Anatomia de La GallinaDocument28 pagesAnatomia de La Gallinavictor alpirezNo ratings yet
- Expo. Enfermedades ZoonoticasDocument94 pagesExpo. Enfermedades ZoonoticasM Christel CortézNo ratings yet
- Preestómagos rumiantesDocument94 pagesPreestómagos rumiantesnatula23No ratings yet
- Hipotricosis CongénitaDocument17 pagesHipotricosis CongénitaJhofre GuatemalNo ratings yet
- Psicología transpersonal y sus principales autoresDocument1 pagePsicología transpersonal y sus principales autoresMAIRA ALEJANDRA MORALES CORDOBANo ratings yet
- ETOLOGÍADocument17 pagesETOLOGÍAMartita JoNo ratings yet
- Salmonella TiphyDocument4 pagesSalmonella TiphyMario alvarez cruzNo ratings yet
- 2 Unidad2 Tejido GlandularDocument10 pages2 Unidad2 Tejido Glandularluna2702No ratings yet
- Unidad 5 Degradacion de Comp. Org.Document16 pagesUnidad 5 Degradacion de Comp. Org.Erika Sanchez HNo ratings yet
- RT-PCR: Reverse Transcribed PCR para detección de ARNDocument3 pagesRT-PCR: Reverse Transcribed PCR para detección de ARNErika Sanchez HNo ratings yet
- Guion - de Practicas - 2003Document9 pagesGuion - de Practicas - 2003Maria Paz Castillo CaceresNo ratings yet
- Qué Son Las Emociones PDFDocument11 pagesQué Son Las Emociones PDFRul QuirozNo ratings yet
- RT-PCR: Reverse Transcribed PCR para detección de ARNDocument3 pagesRT-PCR: Reverse Transcribed PCR para detección de ARNErika Sanchez HNo ratings yet
- Colores PictóricosDocument1 pageColores PictóricosErika Sanchez HNo ratings yet
- RT-PCR: Reverse Transcribed PCR para detección de ARNDocument3 pagesRT-PCR: Reverse Transcribed PCR para detección de ARNErika Sanchez HNo ratings yet
- Arenas MicologiaMedica 4a Capitulo MuestraDocument17 pagesArenas MicologiaMedica 4a Capitulo MuestraDonny TorresNo ratings yet
- 2.09.07. Listeria Monocytogenes PDFDocument18 pages2.09.07. Listeria Monocytogenes PDFErika Sanchez HNo ratings yet
- Taller Biología MolecularDocument21 pagesTaller Biología MolecularErika Sanchez HNo ratings yet
- Alteraciones microbianas en frutas secas: hongos y bacterias que pueden crecer a bajos valores de actividad de aguaDocument1 pageAlteraciones microbianas en frutas secas: hongos y bacterias que pueden crecer a bajos valores de actividad de aguaErika Sanchez HNo ratings yet
- Protocolo WesternDocument1 pageProtocolo WesternErika Sanchez HNo ratings yet
- Protocolo WesternDocument1 pageProtocolo WesternErika Sanchez HNo ratings yet
- Manual CTO - Cardiologia y Cirugia CardiovascularDocument84 pagesManual CTO - Cardiologia y Cirugia CardiovascularMey AcNo ratings yet
- Huesos y MusculosDocument26 pagesHuesos y MusculosJose RomaniNo ratings yet
- Fisiologia de La ContraccionDocument25 pagesFisiologia de La ContraccionAna Lilyam Rios BayonaNo ratings yet
- Matriz Sistema MuscularDocument10 pagesMatriz Sistema MuscularPatiño YessicaNo ratings yet
- D 0036501Document289 pagesD 0036501Maria de DiegoNo ratings yet
- NeurofisiologíaDocument4 pagesNeurofisiologíaXavier VerducciNo ratings yet
- Tzo 051Document183 pagesTzo 051Victorino KjNo ratings yet
- Músculos VoluntariosDocument4 pagesMúsculos VoluntariosBryan CordobaNo ratings yet
- SM. 5 Bases Moleculares de La Contraccion MuscularDocument15 pagesSM. 5 Bases Moleculares de La Contraccion MuscularMario Camilo ColladoNo ratings yet
- Banco de Preguntas 1 P BiomecànicaDocument6 pagesBanco de Preguntas 1 P BiomecànicaRamiro QMNo ratings yet
- Informe M. EstriadoDocument33 pagesInforme M. EstriadodannymalcacamposNo ratings yet
- Miologia de TodoDocument13 pagesMiologia de Todo黒心No ratings yet
- Informe BCM - ProteínasDocument26 pagesInforme BCM - ProteínasIan Gabriel Pérez DintransNo ratings yet
- Flexibilidad PDFDocument108 pagesFlexibilidad PDFCharlotte Francisca Valenzuela RodriguezNo ratings yet
- Exames e Repasos de Histologia (No Completo)Document18 pagesExames e Repasos de Histologia (No Completo)Guido Walter Vargas ApazaNo ratings yet
- Carnicos IIDocument166 pagesCarnicos IIAgustin Pazmiño TorresNo ratings yet
- Conversión Del Músculo en CarneDocument13 pagesConversión Del Músculo en Carneamarlun446No ratings yet
- Contracción Muscular - GuytonDocument9 pagesContracción Muscular - GuytonSanty LopezNo ratings yet
- SISTEMA LOCOMOTORDocument7 pagesSISTEMA LOCOMOTORDUVAN ALEXANDER ADRADA MOSQUERANo ratings yet
- Citoesqueleto PDFDocument20 pagesCitoesqueleto PDFIsidoraNo ratings yet
- Sistema MuscularDocument7 pagesSistema MuscularERIKANo ratings yet
- Apunte Tejido MuscularDocument74 pagesApunte Tejido MuscularGael GilNo ratings yet
- Proceso de Contracion MuscularDocument41 pagesProceso de Contracion Muscularjose reinerio castañeda avila100% (1)
- Unidad 2 Soporte Y MOVIMIENTOSDocument174 pagesUnidad 2 Soporte Y MOVIMIENTOSGuadalupe FloresNo ratings yet
- Entrenamiento Ambientes ExtremosDocument223 pagesEntrenamiento Ambientes ExtremosSergio Xepu100% (1)
- CitoesqueletoDocument45 pagesCitoesqueletoCarlos GiraldoNo ratings yet
- Cardiología y Cirugía CardiovascularDocument31 pagesCardiología y Cirugía Cardiovascularshur mirNo ratings yet
- Resumen HistologiaDocument34 pagesResumen HistologiavalentinaNo ratings yet
- Presentación Guía para El Viaje Perfecto Doodle BeigeDocument34 pagesPresentación Guía para El Viaje Perfecto Doodle BeigeMELANY DIANA OQUENDO GUTIERREZNo ratings yet
- Proteinas de La CarneDocument2 pagesProteinas de La CarneJuberly DelgadoNo ratings yet