You might also like
- Regulación de La GlucólisisDocument3 pagesRegulación de La Glucólisisfrancisco lozano gomezNo ratings yet
- Metabolismo de los hidratos de carbonoDocument28 pagesMetabolismo de los hidratos de carbonoFranco FilippoliNo ratings yet
- Metabolismo Del GlucógenoDocument10 pagesMetabolismo Del GlucógenobiologiaibericaNo ratings yet
- GlucogenolisisDocument7 pagesGlucogenolisisantonioNo ratings yet
- METABOLISMO GLUCOGENODocument34 pagesMETABOLISMO GLUCOGENOEsmeralda CarreraNo ratings yet
- GlucogénesisDocument4 pagesGlucogénesisBryan CorderoNo ratings yet
- Ciclo de Cori: la circulación de glucosa y lactato entre músculo y hígadoDocument7 pagesCiclo de Cori: la circulación de glucosa y lactato entre músculo y hígadoJeanette PaulinaNo ratings yet
- Metabolismo de Otras HexosasDocument4 pagesMetabolismo de Otras HexosasMaria Zimbrelo Alcarria0% (1)
- Vías metabólicas de la glucosaDocument3 pagesVías metabólicas de la glucosaRebeca RioNo ratings yet
- La Insulina y Sus Efectos MetabólicosDocument12 pagesLa Insulina y Sus Efectos MetabólicosOscar Vasquez CorreaNo ratings yet
- Metabolismo Del GlucogenoDocument30 pagesMetabolismo Del GlucogenoMAURICIO AUGUSTO ORELLANA CARDOZO100% (1)
- Páncreas y diabetes: funciones de la insulina y el glucagónDocument8 pagesPáncreas y diabetes: funciones de la insulina y el glucagónStephania Cadena JimenezNo ratings yet
- Ejercicio N°10 - Bioquímica - OCWDocument4 pagesEjercicio N°10 - Bioquímica - OCWYan HernándezNo ratings yet
- Proteína Visceral y EsqueléticaDocument7 pagesProteína Visceral y EsqueléticaNicole RamirezNo ratings yet
- 001-Digestión y Absorción de LípidosDocument27 pages001-Digestión y Absorción de LípidosGreissy Campos Cabrera100% (1)
- Taller GlucólisisDocument2 pagesTaller GlucólisisCarlos Riofrio0% (1)
- Anabolismo de ProteínasDocument11 pagesAnabolismo de ProteínasSEPALABONo ratings yet
- Von Gierke, El InformeDocument9 pagesVon Gierke, El InformeFrancisco Javier Madrid Ruiz100% (3)
- Cuestionario 11Document2 pagesCuestionario 11Mayker PanibraNo ratings yet
- La Secreción de La InsulinaDocument5 pagesLa Secreción de La Insulinaeloydc17No ratings yet
- Gluconeogénesis en La Corteza RenalDocument2 pagesGluconeogénesis en La Corteza RenalNicolas Andres Santiago Becerra100% (2)
- Metabolismo de La GlucosaDocument15 pagesMetabolismo de La GlucosaDayana EstefaniaNo ratings yet
- Funciones Metabólicas Del HígadoDocument3 pagesFunciones Metabólicas Del HígadoEliana Cadena100% (1)
- Las 10 Reacciones de La GlucólisisDocument2 pagesLas 10 Reacciones de La Glucólisisjesus75% (4)
- NEFRONADocument6 pagesNEFRONALuis Fernando BCNo ratings yet
- CuestionarioDocument5 pagesCuestionarioMaríaGabrielaOrellanaNo ratings yet
- Bioquimica de La Diabetes 2Document16 pagesBioquimica de La Diabetes 2AdonayDecoraciones50% (2)
- w4 Clase Bioca Cetogenesis Cetolisis Xenob 20052020Document32 pagesw4 Clase Bioca Cetogenesis Cetolisis Xenob 20052020Darreem Steve Villarreal PutnamNo ratings yet
- Ciclo Del Ácido TricarboxílicoDocument32 pagesCiclo Del Ácido TricarboxílicoDenissLopez100% (1)
- GluconeogénesisDocument1 pageGluconeogénesisLópez chente López Ángel de JesúsNo ratings yet
- Los carbohidratos: fuente de energía y componentes estructuralesDocument4 pagesLos carbohidratos: fuente de energía y componentes estructuralesNicole AndradeNo ratings yet
- Metabolismo de Otros Azúcares ImportantesDocument4 pagesMetabolismo de Otros Azúcares ImportantesAnthony QuevedoNo ratings yet
- Informe Bioquimica 5Document3 pagesInforme Bioquimica 5Cesar Del Aguila Matta67% (6)
- Líquidos corporales: composición, regulación y trastornosDocument5 pagesLíquidos corporales: composición, regulación y trastornosJaime Junior100% (1)
- La Ruta de Las Pentosas FosfatoDocument4 pagesLa Ruta de Las Pentosas FosfatoMiguel MuñozNo ratings yet
- Glucólisis: La ruta universal para extraer energía de la glucosaDocument21 pagesGlucólisis: La ruta universal para extraer energía de la glucosatatahimys0% (1)
- Descarboxilacion OxidativaDocument34 pagesDescarboxilacion Oxidativaelvismartha50% (2)
- Glucólisis, Gluconeogénesis y Ruta de Las Pentosas FosfatoDocument26 pagesGlucólisis, Gluconeogénesis y Ruta de Las Pentosas FosfatoMarcelo Kim100% (1)
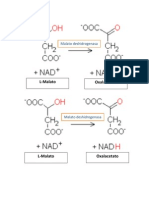
- La Enzima Malato DeshidrogenasaDocument2 pagesLa Enzima Malato DeshidrogenasaKaren Torres JuárezNo ratings yet
- Via de Las PentosasDocument25 pagesVia de Las PentosasHKlaudyNo ratings yet
- Mecanismos de Regulación EnzimáticaDocument2 pagesMecanismos de Regulación EnzimáticaRobert Isaias Mendoza CardenasNo ratings yet
- Síntesis de Ácidos GrasosDocument16 pagesSíntesis de Ácidos GrasosMona Gallegos100% (1)
- Lanza Der AsDocument3 pagesLanza Der AsJose RicardoNo ratings yet
- Isiologia Del Metabolismo de La Proteinas PDFDocument19 pagesIsiologia Del Metabolismo de La Proteinas PDFRosario Laguna100% (1)
- Anabolismo de Carbohidratos 3146Document35 pagesAnabolismo de Carbohidratos 3146Vale MartinezNo ratings yet
- Digestion y Absorcion de LipidosDocument14 pagesDigestion y Absorcion de LipidosHeber Barboza FustamanteNo ratings yet
- Metabolismo eritrocito glucosa ATP protege hemoglobinaDocument11 pagesMetabolismo eritrocito glucosa ATP protege hemoglobinaJGQM08No ratings yet
- Rutas MetabolicasDocument30 pagesRutas Metabolicasamilcar torresNo ratings yet
- Biosíntesis de Carbohidratos - BIOQUIMICA II U2Document13 pagesBiosíntesis de Carbohidratos - BIOQUIMICA II U2Bernardo Morales0% (1)
- Identificación de la glucólisis y fermentaciónDocument11 pagesIdentificación de la glucólisis y fermentaciónyohana tabares romanNo ratings yet
- Efecto de La Insulina en El MúsculoDocument7 pagesEfecto de La Insulina en El MúsculoTania Belen GarciaNo ratings yet
- Digestión y absorción de ácidos nucleicosDocument5 pagesDigestión y absorción de ácidos nucleicosfranklin ruiz100% (1)
- Componentes de La Cadena RespiratoriaDocument5 pagesComponentes de La Cadena RespiratoriaHéctor De La Cruz BenitesNo ratings yet
- Oxidacion de Ácidos GrasosDocument1 pageOxidacion de Ácidos Grasosagustin zarate cruzNo ratings yet
- Cuestionario - Tema 10Document5 pagesCuestionario - Tema 10María del Carmen Ramírez SalinasNo ratings yet
- Beta oxidación de ácidos grasos: proceso catabólico por ciclos de 4 reaccionesDocument16 pagesBeta oxidación de ácidos grasos: proceso catabólico por ciclos de 4 reaccionesshirley100% (1)
- Glucolisis 21645 PDFDocument77 pagesGlucolisis 21645 PDFShärlÿ ContrerasNo ratings yet
- LIPOLISISDocument8 pagesLIPOLISISScarlette OlivoNo ratings yet
- 2Document8 pages2Alba Montes FernandezNo ratings yet
- Genero y Cambio Climatico, 2010Document23 pagesGenero y Cambio Climatico, 2010David Morales NuñezNo ratings yet
- El Papel de La Mujer en El Combate Al Cambio ClimaticoDocument9 pagesEl Papel de La Mujer en El Combate Al Cambio ClimaticoLorik90No ratings yet
- Producción de glucoamilasa por Aspergillus nigerDocument10 pagesProducción de glucoamilasa por Aspergillus nigerLorik90No ratings yet
- Ensayo Sobre Transformación Genética en PlantasDocument2 pagesEnsayo Sobre Transformación Genética en PlantasLorik90No ratings yet
- Estrategia para El Cambio ClimáticoDocument13 pagesEstrategia para El Cambio ClimáticoLorik90No ratings yet
- Ensayo Sobre Aplicaciones Del CTV para La Conservación Del GermoplasmaDocument2 pagesEnsayo Sobre Aplicaciones Del CTV para La Conservación Del GermoplasmaLorik90100% (1)
- Calentamiento Global y Cambio ClimaticoDocument10 pagesCalentamiento Global y Cambio ClimaticoLorik90No ratings yet
- Gases de Efecto Invernadero (Gei)Document9 pagesGases de Efecto Invernadero (Gei)Lorik90No ratings yet
- EL CAMBIO CLIMÁTICO. El Día Que Me Cambió El ClimaDocument47 pagesEL CAMBIO CLIMÁTICO. El Día Que Me Cambió El ClimaJorge AzcorraNo ratings yet
- Crustáceos Con Potencial BiotecnológicoDocument10 pagesCrustáceos Con Potencial BiotecnológicoLorik90No ratings yet
- Escalas Termometricas Absoluta y RelativaDocument1 pageEscalas Termometricas Absoluta y RelativaLorik90No ratings yet
- Via GlucoliticaDocument4 pagesVia GlucoliticaLorik90No ratings yet
- Alimentos Organicos, Moda o NecesidadDocument30 pagesAlimentos Organicos, Moda o NecesidadLorik90No ratings yet
- Alimentos TransgénicosDocument4 pagesAlimentos TransgénicosLorik90No ratings yet
- CrustaceosDocument4 pagesCrustaceosLorik90No ratings yet
- Biotecnología en CrustáceosDocument2 pagesBiotecnología en CrustáceosLorik90No ratings yet
- Grasas TransDocument1 pageGrasas TransLorik90No ratings yet
- Las EnzimasDocument4 pagesLas EnzimasLorik90No ratings yet
- Biotecnología Marina Aplicada A La FarmacologíaDocument3 pagesBiotecnología Marina Aplicada A La FarmacologíaLorik90No ratings yet
- Ámbitos de Aplicación de La Biotecnología MarinaDocument3 pagesÁmbitos de Aplicación de La Biotecnología MarinaLorik90No ratings yet
- BIORREACTORESDocument5 pagesBIORREACTORESLorik90No ratings yet
- Via GlucoliticaDocument4 pagesVia GlucoliticaLorik90No ratings yet
- Cultivo de CobiaDocument3 pagesCultivo de CobiaLorik90No ratings yet
- Almidones ModificadosDocument2 pagesAlmidones ModificadosLorik90No ratings yet
- NITRATOSDocument4 pagesNITRATOSLorik90No ratings yet
- Depuracion de OstionesDocument3 pagesDepuracion de OstionesLorik90No ratings yet
- CARBOHIDRATOSDocument2 pagesCARBOHIDRATOSLorik90No ratings yet
- COENZIMASDocument3 pagesCOENZIMASLorik90No ratings yet
- Conceptos y aplicaciones de la biotecnología marinaDocument6 pagesConceptos y aplicaciones de la biotecnología marinaLorik90No ratings yet
- TEMA 2 GAMETOGENESIS DRA. SANTISTEBANDocument20 pagesTEMA 2 GAMETOGENESIS DRA. SANTISTEBANaaronruizlijeron2024No ratings yet
- Práctica Tejido SanguíneoDocument4 pagesPráctica Tejido SanguíneoMónica CantúNo ratings yet
- Biologia CelularDocument45 pagesBiologia CelularRocío PonceNo ratings yet
- Mitosis CebollaDocument2 pagesMitosis Cebolladiegonzalez25100% (1)
- Estructura CelularDocument8 pagesEstructura CelularKelly GmzNo ratings yet
- Informe Final Laboratorio Biología-1Document31 pagesInforme Final Laboratorio Biología-1Angel Elvith Aguilar92% (12)
- 2a Metabolismo Microbiano II 22Document36 pages2a Metabolismo Microbiano II 22Ambar DiazNo ratings yet
- AA - Test Histología AnimalDocument4 pagesAA - Test Histología AnimalLaura ClementeNo ratings yet
- Funcionamiento Celular y EmbriogénesisDocument14 pagesFuncionamiento Celular y EmbriogénesisSofy Uco100% (1)
- Espermatogenesis y OvogenesisDocument17 pagesEspermatogenesis y OvogenesisBlake Agustin PoncinNo ratings yet
- TemasDocument18 pagesTemasXtlv SaykoNo ratings yet
- Niveles de Organización. EjerciciosDocument1 pageNiveles de Organización. EjerciciosMaricieloNo ratings yet
- Ciencias Naturales: Grado Séptimo Taller de RefuerzoDocument17 pagesCiencias Naturales: Grado Séptimo Taller de RefuerzoCésar Andrés100% (1)
- Taller Metabolismo en ClaseDocument6 pagesTaller Metabolismo en ClaseAshly Romero JiménezNo ratings yet
- ANTÍGENOS. Estructura Tipos e ImportanciaDocument30 pagesANTÍGENOS. Estructura Tipos e ImportanciaChristian Blanqviazvl0% (1)
- Hepatitis Por ToxicosDocument7 pagesHepatitis Por Toxicosmuter007100% (1)
- Taller #1 - La Celula - 10Document37 pagesTaller #1 - La Celula - 10Alexa HernándezNo ratings yet
- Celula VegetalDocument6 pagesCelula VegetalNaydu JaimesNo ratings yet
- Receptores de Reconocimiento de PatronDocument8 pagesReceptores de Reconocimiento de PatronJesús FraustoNo ratings yet
- Membrana PlasmáticaDocument6 pagesMembrana PlasmáticaNestorNo ratings yet
- MHCDocument69 pagesMHCfrank208647259No ratings yet
- HEMOSTASIADocument40 pagesHEMOSTASIAjosephNo ratings yet
- Taller BiologíaDocument3 pagesTaller BiologíaAdriana PanquevaNo ratings yet
- Mapa Mental Unión A ProteínasDocument1 pageMapa Mental Unión A ProteínasSANDRA ABIGAIL ORTEGA MOLINARNo ratings yet
- Dra Carmen Fisiologia 1 Era ClaseDocument49 pagesDra Carmen Fisiologia 1 Era ClaseCarlos FernandoNo ratings yet
- BHE Mapa MentalDocument2 pagesBHE Mapa MentalAreli Marisol Guzman100% (1)
- Resumen Bioquimica DMDDocument2 pagesResumen Bioquimica DMDGIOVANNA BARROSNo ratings yet
- CelulasDocument8 pagesCelulasAra VeraNo ratings yet
- Taller Oxidación Biológica Final-2020-1Document1 pageTaller Oxidación Biológica Final-2020-1Fabio Fajardo67% (3)
- Quimica FinlDocument6 pagesQuimica FinlInes maria Huaqui caleroNo ratings yet