You might also like
- Biochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967From EverandBiochemical Factors Concerned in the Functional Activity of the Nervous System: First International Meeting of the International Society for Neurochemistry, Strasbourg, 1967D. RichterNo ratings yet
- Biochemical and Pharmacological Roles of Adenosylmethionine and the Central Nervous System: Proceedings of an International Round Table on Adenosylmethionine and the Central Nervous System, Naples, Italy, May 1978From EverandBiochemical and Pharmacological Roles of Adenosylmethionine and the Central Nervous System: Proceedings of an International Round Table on Adenosylmethionine and the Central Nervous System, Naples, Italy, May 1978Vincenzo ZappiaNo ratings yet
- Role of Reactive Oxygen Species in p53 Activation During Cisplatin-Induced Apoptosis of Rat Mesangial CellsDocument7 pagesRole of Reactive Oxygen Species in p53 Activation During Cisplatin-Induced Apoptosis of Rat Mesangial CellsPandji 'Utuh'No ratings yet
- 1 FullDocument9 pages1 FullFauziana NurhanisahNo ratings yet
- Said AhmedDocument19 pagesSaid Ahmedahmed mahdiNo ratings yet
- SJ BJP 0706338Document8 pagesSJ BJP 0706338Rogério HitsugayaNo ratings yet
- Reisser2002 Ic50Document4 pagesReisser2002 Ic50Jesús Rafael Méndez NateraNo ratings yet
- Monitoring of ATP Levels in Red Blood Cells and T Cells of Healthy and Ill Subjects and The Effects of Age On Mitochondrial PotentialDocument9 pagesMonitoring of ATP Levels in Red Blood Cells and T Cells of Healthy and Ill Subjects and The Effects of Age On Mitochondrial PotentialHossam ArafaNo ratings yet
- Kaemferol and Abeta PDFDocument5 pagesKaemferol and Abeta PDFKapil SoniNo ratings yet
- Science 273 5280 1380Document5 pagesScience 273 5280 1380Arslan HamidNo ratings yet
- Beta Sitosterol y GSHDocument7 pagesBeta Sitosterol y GSHEridani OlmosNo ratings yet
- PDFDocument11 pagesPDFAstridz Monsteqszia SilvanaNo ratings yet
- Mikula 2006Document12 pagesMikula 2006julia rossoNo ratings yet
- Ajplung 1993 265 3 l286Document7 pagesAjplung 1993 265 3 l286moussaouiNo ratings yet
- Cellular Antioxidant Effects of Atorvastatin in Vitro and in VivoDocument6 pagesCellular Antioxidant Effects of Atorvastatin in Vitro and in VivoantonopoulosalNo ratings yet
- Nuclear Diameter in The Anterior Pituitary Gland of The RatDocument5 pagesNuclear Diameter in The Anterior Pituitary Gland of The RatLonkesNo ratings yet
- Ligustrazine Phosphate Ethosomes For Treatment of Alzheimer's Disease, in Vitro and in Animal Model StudiesDocument8 pagesLigustrazine Phosphate Ethosomes For Treatment of Alzheimer's Disease, in Vitro and in Animal Model Studiesebooks list1No ratings yet
- NT 5mmbleed 1Document6 pagesNT 5mmbleed 1Diana MihaiNo ratings yet
- Pinealon Increases Cell Viability by Suppression of Free Radical Levels and Activating Proliferative Processes.Document8 pagesPinealon Increases Cell Viability by Suppression of Free Radical Levels and Activating Proliferative Processes.scribdNo ratings yet
- TMP AE80Document6 pagesTMP AE80FrontiersNo ratings yet
- Sloane1981 - Science - CathepsinsDocument4 pagesSloane1981 - Science - CathepsinsPilar AufrastoNo ratings yet
- Circulating Hormone Adrenomedullin and Its Binding Protein Protect Neural Cells From Hypoxia-Induced ApoptosisDocument29 pagesCirculating Hormone Adrenomedullin and Its Binding Protein Protect Neural Cells From Hypoxia-Induced ApoptosisShieldNo ratings yet
- Antioxidant Treatment Induces Hyperactivation of The HPA Axis by Upregulating ACTH Receptor in The Adrenal and Downregulating Glucocorticoid Receptors in The PituitaryDocument13 pagesAntioxidant Treatment Induces Hyperactivation of The HPA Axis by Upregulating ACTH Receptor in The Adrenal and Downregulating Glucocorticoid Receptors in The PituitaryVũ Đoàn Ngọc HưngNo ratings yet
- Ja8b11449 Si 001Document29 pagesJa8b11449 Si 001Angel Ernesto Perales RochaNo ratings yet
- Endotelial Pericites - Cui2006Document10 pagesEndotelial Pericites - Cui2006Lilo FrancoisNo ratings yet
- Malate-aspartate shuttle mediates antioxidation of neural cellsDocument6 pagesMalate-aspartate shuttle mediates antioxidation of neural cellsZevannayasmineNo ratings yet
- Papaer ArthurDocument11 pagesPapaer ArthurrrresendeNo ratings yet
- Journal 6Document7 pagesJournal 6Claudia JessicaNo ratings yet
- Nitric Oxide, CGMP, and Hormone Regulation of Active Sodium TransportDocument6 pagesNitric Oxide, CGMP, and Hormone Regulation of Active Sodium TransportMichael Daley100% (1)
- Synthesis of hemoglobin in a cell-free systemDocument8 pagesSynthesis of hemoglobin in a cell-free systemDuaa ShamiehNo ratings yet
- Losartan Reduces NF-κB Activation by Small Dense LDLDocument4 pagesLosartan Reduces NF-κB Activation by Small Dense LDLaditya_permana77No ratings yet
- English 2Document7 pagesEnglish 2Jc GaldosNo ratings yet
- jlrM062752 PDFDocument13 pagesjlrM062752 PDFTheodora TeddyNo ratings yet
- Nchembio 412-S1Document21 pagesNchembio 412-S1FranciscoNo ratings yet
- Anthocyanins Induce Cholesterol Efflux From Mouse Peritoneal MacrophagesDocument10 pagesAnthocyanins Induce Cholesterol Efflux From Mouse Peritoneal MacrophagesTri Anna FitrianiNo ratings yet
- Protective Effects of Lagerstroemia Speciosa On 3-Morpholinosydnonimine (SIN-1) - Induced Oxidative Stress in HIT-T15 Pancreatic CellsDocument6 pagesProtective Effects of Lagerstroemia Speciosa On 3-Morpholinosydnonimine (SIN-1) - Induced Oxidative Stress in HIT-T15 Pancreatic CellsCurieThamerNo ratings yet
- Effect of Quercetin On Paraoxonase 1 Activity - Studies in Cultured Cells, Mice and HumansDocument7 pagesEffect of Quercetin On Paraoxonase 1 Activity - Studies in Cultured Cells, Mice and Humansanisa silviNo ratings yet
- Z. Yan Et Al - Coordinated Expression of Muscarininc Receptor Messenger RNAs in Straital Medium Spiny NeuronsDocument8 pagesZ. Yan Et Al - Coordinated Expression of Muscarininc Receptor Messenger RNAs in Straital Medium Spiny NeuronsJunmajNo ratings yet
- Adenylyl Cyclase in The Heart: An Enzymocytochemical and Immunocytochemical ApproachDocument6 pagesAdenylyl Cyclase in The Heart: An Enzymocytochemical and Immunocytochemical ApproachJoNo ratings yet
- 34 77lllllllllllllDocument5 pages34 77lllllllllllllJean Pierre Chastre LuzaNo ratings yet
- Cytotoxic Effects of Repin, A Principal Sesquiterpene Lactone of Russian KnapweedDocument8 pagesCytotoxic Effects of Repin, A Principal Sesquiterpene Lactone of Russian KnapweedcthjgfytvfnoNo ratings yet
- N Gonore 2Document8 pagesN Gonore 2qurrotun faizahNo ratings yet
- Reduced Homocysteine-Thiolactonase Activity in Alzheimer's DiseaseDocument7 pagesReduced Homocysteine-Thiolactonase Activity in Alzheimer's DiseaseAlfonso De la FuenteNo ratings yet
- Protective Effect of Antioxidant Enzymes Against Drug Cytotoxicity in Mcf-7 CellsDocument3 pagesProtective Effect of Antioxidant Enzymes Against Drug Cytotoxicity in Mcf-7 CellsVidhi AdilNo ratings yet
- DNA Repair Protein Involved in Heart and Blood DevelopmentDocument12 pagesDNA Repair Protein Involved in Heart and Blood DevelopmentSol Jumaide WerbleNo ratings yet
- Spermine Mediated Negative Inotropic Effect via ATP ReleaseDocument5 pagesSpermine Mediated Negative Inotropic Effect via ATP Releaseuaslp_20988No ratings yet
- Acido Ursolico NNDocument5 pagesAcido Ursolico NNDIANA PATRICIA MOLANO PEREZNo ratings yet
- Glycolysis-Deficient: Studies Cells That Production of TumorDocument5 pagesGlycolysis-Deficient: Studies Cells That Production of TumorFabian BalderrabanoNo ratings yet
- PIIS0021925819685212Document6 pagesPIIS0021925819685212windfragNo ratings yet
- New ResearchDocument6 pagesNew ResearchChloe MirandaNo ratings yet
- UntitledDocument7 pagesUntitledTuấn Nguyen AnhNo ratings yet
- Ribosome-Inactivating Protein and Apoptosis: Abrin Causes Cell Death Via Mitochondrial Pathway in Jurkat CellsDocument8 pagesRibosome-Inactivating Protein and Apoptosis: Abrin Causes Cell Death Via Mitochondrial Pathway in Jurkat CellsKusuma WardaniNo ratings yet
- TMP 857 FDocument11 pagesTMP 857 FFrontiersNo ratings yet
- PNAS 2000 Budd 6161 6Document6 pagesPNAS 2000 Budd 6161 6Aulia MuhammadNo ratings yet
- Calorie RestrictionDocument18 pagesCalorie RestrictionJuan Felipe QuinteroNo ratings yet
- Potent Vasoconstrictor Kisspeptin-10 Induces Atherosclerotic Plaque Progression and Instability: Reversal by Its Receptor GPR54 AntagonistDocument20 pagesPotent Vasoconstrictor Kisspeptin-10 Induces Atherosclerotic Plaque Progression and Instability: Reversal by Its Receptor GPR54 AntagonistÁlex GómezNo ratings yet
- Hildegarde Esther Allen: City of Medical AND Division OF OFDocument7 pagesHildegarde Esther Allen: City of Medical AND Division OF OFJulien Patrick CebrianNo ratings yet
- TJPS 0 0 enDocument18 pagesTJPS 0 0 enOlukunle DamolaNo ratings yet
- Oxidative Stress Is The Primary Event: Effects of Ethanol Consumption in BrainDocument6 pagesOxidative Stress Is The Primary Event: Effects of Ethanol Consumption in BrainMokhammad Faisol AbdullahNo ratings yet
- Soluble A Peptide Increases Excitatory Neurotransmission and Induces Epileptiform Activity in Hippocampal NeuronsDocument15 pagesSoluble A Peptide Increases Excitatory Neurotransmission and Induces Epileptiform Activity in Hippocampal NeuronsClaudia Alejandra Lopez ToroNo ratings yet
- Abbas, Al-Arini, Meshikhes - 2014 - Gastric Autonomic Nerve Tumour A Rare Gastric Tumour PDFDocument2 pagesAbbas, Al-Arini, Meshikhes - 2014 - Gastric Autonomic Nerve Tumour A Rare Gastric Tumour PDFflashjetNo ratings yet
- Abbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientDocument4 pagesAbbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientflashjetNo ratings yet
- Nana Et Al. - 2014 - Left Renal Vein Thrombosis A Rare Cause of Acute Scrotal Pain PDFDocument2 pagesNana Et Al. - 2014 - Left Renal Vein Thrombosis A Rare Cause of Acute Scrotal Pain PDFflashjetNo ratings yet
- Abbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewDocument5 pagesAbbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewflashjetNo ratings yet
- Abbas Et Al. - 2020 - Maneuvering The Management of A Rare Case of Primary Undifferentiated Cardiac Sarcoma PDFDocument5 pagesAbbas Et Al. - 2020 - Maneuvering The Management of A Rare Case of Primary Undifferentiated Cardiac Sarcoma PDFflashjetNo ratings yet
- Abbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewDocument5 pagesAbbas Et Al. - 2016 - Unusual Complication of Pituitary Macroadenoma A Case Report and ReviewflashjetNo ratings yet
- Abangah, Rashidbeygi - 2013 - Osler-Weber-Rendu SyndromeDocument2 pagesAbangah, Rashidbeygi - 2013 - Osler-Weber-Rendu SyndromeflashjetNo ratings yet
- Namburar Et Al. - 2018 - Waste Generated During Glaucoma Surgery A Comparison of Two Global Facilities PDFDocument4 pagesNamburar Et Al. - 2018 - Waste Generated During Glaucoma Surgery A Comparison of Two Global Facilities PDFflashjetNo ratings yet
- Nascimento Et Al. - 2012 - Adrenoleukodystrophy A Forgotten Diagnosis in Children With Primary Addison' S DiseaseDocument5 pagesNascimento Et Al. - 2012 - Adrenoleukodystrophy A Forgotten Diagnosis in Children With Primary Addison' S DiseaseflashjetNo ratings yet
- Nascimento Et Al. - 2013 - Asymptomatic Anomalous Left Anterior Descending Artery Arising From The Right Coronary Artery With A Rare AntDocument3 pagesNascimento Et Al. - 2013 - Asymptomatic Anomalous Left Anterior Descending Artery Arising From The Right Coronary Artery With A Rare AntflashjetNo ratings yet
- Narula, Nangia - 2014 - Use of An Oscillatory PEP Device To Enhance Bronchial Hygiene in A Patient of post-H1NI Pneumonia and Acute Resp PDFDocument3 pagesNarula, Nangia - 2014 - Use of An Oscillatory PEP Device To Enhance Bronchial Hygiene in A Patient of post-H1NI Pneumonia and Acute Resp PDFflashjetNo ratings yet
- Nascimento Et Al. - 2012 - Bloody Nipple Discharge in Infancy - Report of Two CasesDocument3 pagesNascimento Et Al. - 2012 - Bloody Nipple Discharge in Infancy - Report of Two CasesflashjetNo ratings yet
- Abbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientDocument4 pagesAbbas Et Al. - 2013 - Posterior Reversible Encephalopathy Syndrome After Bevacizumab Therapy in A Normotensive PatientflashjetNo ratings yet
- Nampei Et Al. - 2020 - Comparison of Ocular Surface Squamous Neoplasia and Pterygium Using Anterior Segment Optical Coherence Tomography PDFDocument4 pagesNampei Et Al. - 2020 - Comparison of Ocular Surface Squamous Neoplasia and Pterygium Using Anterior Segment Optical Coherence Tomography PDFflashjetNo ratings yet
- Nasrat, Al-Alwan - 2012 - Medical Intervention in A Constitutionally Tall ChildDocument3 pagesNasrat, Al-Alwan - 2012 - Medical Intervention in A Constitutionally Tall ChildflashjetNo ratings yet
- Nasser Et Al. - 2013 - Use of Transoesophageal Echocardiography in Endovascular Stenting For Superior Vena Cava SyndromeDocument3 pagesNasser Et Al. - 2013 - Use of Transoesophageal Echocardiography in Endovascular Stenting For Superior Vena Cava SyndromeflashjetNo ratings yet
- Nascimento Et Al. - 2013 - Wearable Cardioverter Defibrillator in Stress Cardiomyopathy and Cardiac ArrestDocument4 pagesNascimento Et Al. - 2013 - Wearable Cardioverter Defibrillator in Stress Cardiomyopathy and Cardiac ArrestflashjetNo ratings yet
- Nassani Et Al. - 2017 - Foreign Body Penetration Through Jejunal Loops Causing Renal Artery Thrombosis and Renal InfarctDocument3 pagesNassani Et Al. - 2017 - Foreign Body Penetration Through Jejunal Loops Causing Renal Artery Thrombosis and Renal InfarctflashjetNo ratings yet
- Stroke and Transient Ischaemic Attack in Over 16s Diagnosis and Initial Management PDF 975574676437Document29 pagesStroke and Transient Ischaemic Attack in Over 16s Diagnosis and Initial Management PDF 975574676437MaryamNo ratings yet
- Nassar, Abdul-Jawad, Barnes - 2018 - Hepatic Failure Due To Cholestatic Hepatitis C in An Immunosuppressed Patient Treated With ElbasvirDocument3 pagesNassar, Abdul-Jawad, Barnes - 2018 - Hepatic Failure Due To Cholestatic Hepatitis C in An Immunosuppressed Patient Treated With ElbasvirflashjetNo ratings yet
- Natrella Et Al. - 2012 - Rare Disease Supratentorial Neurenteric Cyst Associated With A Intraparenchymal SubependymomaDocument6 pagesNatrella Et Al. - 2012 - Rare Disease Supratentorial Neurenteric Cyst Associated With A Intraparenchymal SubependymomaflashjetNo ratings yet
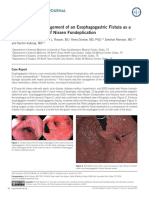
- Naveed Et Al. - 2015 - Diagnosis and Management of An Esophagogastric Fistula As A Rare Complication of Nissen FundoplicationDocument2 pagesNaveed Et Al. - 2015 - Diagnosis and Management of An Esophagogastric Fistula As A Rare Complication of Nissen FundoplicationflashjetNo ratings yet
- Naveen Et Al. - 2014 - Alternatives For Restoration of A Hemisected Mandibular MolarDocument4 pagesNaveen Et Al. - 2014 - Alternatives For Restoration of A Hemisected Mandibular MolarflashjetNo ratings yet
- Nguyen Et Al. - 2020 - Human Leukocyte Antigen Susceptibility Map For Severe Acute Respiratory Syndrome Coronavirus 2Document36 pagesNguyen Et Al. - 2020 - Human Leukocyte Antigen Susceptibility Map For Severe Acute Respiratory Syndrome Coronavirus 2flashjetNo ratings yet
- Idiopathic Stroke After Syndromic and Neuromuscular Scoliosis Surgery: A Case Report and Literature ReviewDocument7 pagesIdiopathic Stroke After Syndromic and Neuromuscular Scoliosis Surgery: A Case Report and Literature ReviewflashjetNo ratings yet
- Naveen Et Al. - 2013 - Complete Denture With Pharyngeal Bulb ProsthesisDocument4 pagesNaveen Et Al. - 2013 - Complete Denture With Pharyngeal Bulb ProsthesisflashjetNo ratings yet
- Nathwani - 2013 - A Complicated Case of AnticoagulationDocument2 pagesNathwani - 2013 - A Complicated Case of AnticoagulationflashjetNo ratings yet
- Nguyen Et Al. - 2019 - Challenges in Management of Autoimmune Hepatitis With Concurrent Graves ThyrotoxicosisDocument4 pagesNguyen Et Al. - 2019 - Challenges in Management of Autoimmune Hepatitis With Concurrent Graves ThyrotoxicosisflashjetNo ratings yet
- Notes 190915 213814 Ea7 PDFDocument1 pageNotes 190915 213814 Ea7 PDFflashjetNo ratings yet
- Notes 200430 195226 0c2 PDFDocument19 pagesNotes 200430 195226 0c2 PDFflashjetNo ratings yet
- E Thesis Uas DharwadDocument5 pagesE Thesis Uas DharwadLori Head100% (1)
- Storyboard: Video Script and Storyboard On Evaluating Functions LessonDocument2 pagesStoryboard: Video Script and Storyboard On Evaluating Functions LessonIvanhoe BalaroteNo ratings yet
- Divya Shree: ObjectiveDocument2 pagesDivya Shree: Objectivekids montessoriNo ratings yet
- Question Bank (ASP)Document3 pagesQuestion Bank (ASP)Gurleen BabraNo ratings yet
- Bcom MADocument21 pagesBcom MAAbl SasankNo ratings yet
- Mikro Prism Manual English PDFDocument18 pagesMikro Prism Manual English PDFrakeshr2007No ratings yet
- Corporate LendingDocument2 pagesCorporate LendingrubaNo ratings yet
- Freebie The Great Composerslapbookseries ChopinDocument24 pagesFreebie The Great Composerslapbookseries ChopinAnonymous EzNMLt0K4C100% (1)
- Taketo SUZUKIDocument3 pagesTaketo SUZUKIJane DeckerNo ratings yet
- Chapter 22 Managing Personal Communications - Direct and Database Marketing and Personal SellingDocument17 pagesChapter 22 Managing Personal Communications - Direct and Database Marketing and Personal SellingBrian Duela100% (1)
- Carbohydrates ReviewerDocument2 pagesCarbohydrates ReviewerJazer AvellanozaNo ratings yet
- M-Commerce Seminar ReportDocument13 pagesM-Commerce Seminar ReportMahar KumarNo ratings yet
- AYUDA Multi-Words VerbsDocument6 pagesAYUDA Multi-Words VerbsGabriel RodriguezNo ratings yet
- Jewish Standard, December 16, 2016 With Beautiful Beginnings and About Our Children SupplementsDocument120 pagesJewish Standard, December 16, 2016 With Beautiful Beginnings and About Our Children SupplementsNew Jersey Jewish StandardNo ratings yet
- Muscovite Mica Flakes and Mica Powder For Industrial ApplicationsDocument5 pagesMuscovite Mica Flakes and Mica Powder For Industrial ApplicationsAnuradhaPatraNo ratings yet
- Anna Kornbluh - Marxist Film Theory and Fight Club (2019, Bloomsbury Academic)Document195 pagesAnna Kornbluh - Marxist Film Theory and Fight Club (2019, Bloomsbury Academic)Yunus ErdoğanNo ratings yet
- What Are The Difficulties With IELTS Listening Multiple Choice Questions?Document9 pagesWhat Are The Difficulties With IELTS Listening Multiple Choice Questions?leoNo ratings yet
- Richardson & McKenzie 1994 Taxonomy Distribution Australian CephalochordatesDocument17 pagesRichardson & McKenzie 1994 Taxonomy Distribution Australian CephalochordatesAmphipodoNo ratings yet
- UPSC EPFO Previous Year Paper 2017 Questions PDFDocument12 pagesUPSC EPFO Previous Year Paper 2017 Questions PDFPraveenNo ratings yet
- Sinha-Dhanalakshmi2019 Article EvolutionOfRecommenderSystemOv PDFDocument20 pagesSinha-Dhanalakshmi2019 Article EvolutionOfRecommenderSystemOv PDFRui MatosNo ratings yet
- Ashtadhyayi of PaniniDocument19 pagesAshtadhyayi of Paniniall4downloads50% (2)
- Customer Satisfaction Towards Honda Two Wheeler: Presented By: Somil Modi (20152002) BBA-MBA 2015Document9 pagesCustomer Satisfaction Towards Honda Two Wheeler: Presented By: Somil Modi (20152002) BBA-MBA 2015Inayat BaktooNo ratings yet
- Data Compilation TemplateDocument257 pagesData Compilation TemplateemelyseuwaseNo ratings yet
- Madbarz Routines Plan: Strength - AdvancedDocument18 pagesMadbarz Routines Plan: Strength - AdvancedJuan Sebastian AnguloNo ratings yet
- Test Bank For Behavior Modification Principles and Procedures 6th EditionDocument36 pagesTest Bank For Behavior Modification Principles and Procedures 6th Editionempericetagragyj6f8100% (31)
- GW and MM Firmware UpgradesDocument11 pagesGW and MM Firmware UpgradesRohit GuptaNo ratings yet
- Enterprise Poem by Nissim Ezekiel Summary and AnalysisDocument7 pagesEnterprise Poem by Nissim Ezekiel Summary and AnalysisSk BMNo ratings yet
- Cultural Etiquette and Business Practices in FranceDocument15 pagesCultural Etiquette and Business Practices in FranceVicky GuptaNo ratings yet
- İngilizce A1 Seviye FiillerDocument70 pagesİngilizce A1 Seviye FiillerMehmet DursunNo ratings yet
- A Time To KillDocument148 pagesA Time To KillMA IZ0% (2)