You might also like
- Tribal Population in Dahod District of Gujarat: An Evaluation of Socio-Economic ProfileDocument9 pagesTribal Population in Dahod District of Gujarat: An Evaluation of Socio-Economic ProfileIJCIRAS Research PublicationNo ratings yet
- Discussion On Comparison of Naturalism of Rabindra Nath Tagor and Mahatma GandhiDocument3 pagesDiscussion On Comparison of Naturalism of Rabindra Nath Tagor and Mahatma GandhiIJCIRAS Research PublicationNo ratings yet
- Diversity of Ant Species (Hymenoptera: Formicidae) in The Campus of Kongunadu Arts and Science College, Coimbatore District, Tamil Nadu.Document5 pagesDiversity of Ant Species (Hymenoptera: Formicidae) in The Campus of Kongunadu Arts and Science College, Coimbatore District, Tamil Nadu.IJCIRAS Research PublicationNo ratings yet
- Perspective of Human Rights in IslamDocument7 pagesPerspective of Human Rights in IslamIJCIRAS Research PublicationNo ratings yet
- Analysis On Stock Market Prediction Using Machine Learning TechniquesDocument5 pagesAnalysis On Stock Market Prediction Using Machine Learning TechniquesIJCIRAS Research PublicationNo ratings yet
- Access Control Verification System Using Statistical MethodsDocument6 pagesAccess Control Verification System Using Statistical MethodsIJCIRAS Research PublicationNo ratings yet
- Pediatric Research: Challenges and Innovative MeasuresDocument3 pagesPediatric Research: Challenges and Innovative MeasuresIJCIRAS Research PublicationNo ratings yet
- Identifying Kantian Aesthetics: A Select Study of Coleridge's PoetryDocument8 pagesIdentifying Kantian Aesthetics: A Select Study of Coleridge's PoetryIJCIRAS Research PublicationNo ratings yet
- Resilience, Learned Optimism and Self Esteem As Predictors of Psychological Well BeingDocument4 pagesResilience, Learned Optimism and Self Esteem As Predictors of Psychological Well BeingIJCIRAS Research PublicationNo ratings yet
- Path Loss Prediction by Using Rssi ValuesDocument7 pagesPath Loss Prediction by Using Rssi ValuesIJCIRAS Research PublicationNo ratings yet
- Popularity of Marxist School of Thought in African Literature: A Critique.Document7 pagesPopularity of Marxist School of Thought in African Literature: A Critique.IJCIRAS Research PublicationNo ratings yet
- Seismic Response of Reinforced Concrete Building Having Ground Soft StoreyDocument5 pagesSeismic Response of Reinforced Concrete Building Having Ground Soft StoreyIJCIRAS Research PublicationNo ratings yet
- Factors Affecting Ready Mix Concrete OperationDocument4 pagesFactors Affecting Ready Mix Concrete OperationIJCIRAS Research PublicationNo ratings yet
- Comparative Study of Sections Used in Industrial Roof Truss and Portal FrameDocument5 pagesComparative Study of Sections Used in Industrial Roof Truss and Portal FrameIJCIRAS Research PublicationNo ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (400)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
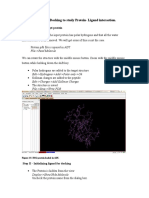
- Molecular Docking Tostudy Protein-Ligand InteractionDocument5 pagesMolecular Docking Tostudy Protein-Ligand InteractionDr. Kaushal Kishor SharmaNo ratings yet
- Boinformatics NewDocument97 pagesBoinformatics NewBob BillsNo ratings yet
- AtikahDocument15 pagesAtikahIka BakarNo ratings yet
- Auto Dock 4.2.6 UserGuideDocument69 pagesAuto Dock 4.2.6 UserGuideBis Chem0% (1)
- Bohacek 1996Document48 pagesBohacek 1996Ronaldo PaxDeorumNo ratings yet
- Ligand-Based+structure-Based ScreeningDocument107 pagesLigand-Based+structure-Based ScreeningManoharNo ratings yet
- Antifungal Activity of Cardiospermum Halicacabum L Sapindaceae Against Trichophyton Rubrum Occurs Through Molecular Interaction With Fungal Hsp90Document10 pagesAntifungal Activity of Cardiospermum Halicacabum L Sapindaceae Against Trichophyton Rubrum Occurs Through Molecular Interaction With Fungal Hsp90abznaim420No ratings yet
- MethodsDocument11 pagesMethodsNguyễn Hiếu HiềnNo ratings yet
- Insilico Study of Curcumin NPDocument20 pagesInsilico Study of Curcumin NPSathish SizzyNo ratings yet
- Computational Intelligence in Medical Informatics PDFDocument105 pagesComputational Intelligence in Medical Informatics PDFHeriberto Aguirre MenesesNo ratings yet
- MVD Manual PDFDocument321 pagesMVD Manual PDFlNo ratings yet
- CCCCCCCCCCCCCCCCC C CCCCCCCCCCCCCCCCCCCCCDocument42 pagesCCCCCCCCCCCCCCCCC C CCCCCCCCCCCCCCCCCCCCCAnkur PandeyNo ratings yet
- In Silico in Drug Discovery TechniquesDocument21 pagesIn Silico in Drug Discovery Techniquesvershaparcha100% (1)
- Docking & Designing Small Molecules Within The Rosetta Code FrameworkDocument64 pagesDocking & Designing Small Molecules Within The Rosetta Code FrameworkGordon LemmonNo ratings yet
- Enzyme Inhibition Studies of AntipyrineDocument10 pagesEnzyme Inhibition Studies of Antipyrineشمس صبيح عبد الرحيمNo ratings yet
- Gold ManualDocument126 pagesGold ManualRamaiyan DhanapalNo ratings yet
- In SilicoDocument8 pagesIn SilicoShafa ShaviraNo ratings yet
- An Overview On Molecular DockingDocument14 pagesAn Overview On Molecular DockingRiya PremNo ratings yet
- Introduction To Pharmacophores in MOEDocument40 pagesIntroduction To Pharmacophores in MOEFatma Fabigha100% (2)
- Medicinal Lab 1Document7 pagesMedicinal Lab 1Adnan RehmatNo ratings yet
- Jurnal Praktikum Kimia Medisinal Kel. 2Document7 pagesJurnal Praktikum Kimia Medisinal Kel. 2AndiNo ratings yet
- Sudipta Baur - Mehebub AlamDocument19 pagesSudipta Baur - Mehebub AlamSudip MajiNo ratings yet
- Bespoke Library Docking For 5-HT Receptor Agonists With Antidepressant ActivityDocument36 pagesBespoke Library Docking For 5-HT Receptor Agonists With Antidepressant Activity别康民No ratings yet
- 8th Sem SyllabusDocument32 pages8th Sem SyllabusDRx Ijajul HussainNo ratings yet
- Immunomodulatory Potential of Bioactive Compounds of Betel LeafDocument14 pagesImmunomodulatory Potential of Bioactive Compounds of Betel LeafHồ KhảiNo ratings yet
- Structural Studies On Docking Selective COX-2 InhibitorsDocument6 pagesStructural Studies On Docking Selective COX-2 InhibitorsFx Hendrick SebastianNo ratings yet
- First Steps in KNIMEDocument19 pagesFirst Steps in KNIMEAldo SotomayorNo ratings yet
- Verdonk 2003Document15 pagesVerdonk 2003AngelCo PetsNo ratings yet
- Gli55 User ManualDocument126 pagesGli55 User Manualcvkkkk1No ratings yet
- Laporan DockingDocument17 pagesLaporan DockingZulfa SeffNo ratings yet