You might also like
- AWS Academy Cloud Architecting Module 04 Student Guide: 200-ACACAD-20-EN-SGDocument89 pagesAWS Academy Cloud Architecting Module 04 Student Guide: 200-ACACAD-20-EN-SGM Rizki AsriyandaNo ratings yet
- ECM PINOUT Chevrolet-Sail PDFDocument2 pagesECM PINOUT Chevrolet-Sail PDFOliver leal90% (10)
- Learning in Neural Models With Complex Dynamics : Michael Stiber SegundoDocument4 pagesLearning in Neural Models With Complex Dynamics : Michael Stiber SegundoHilton SeheresNo ratings yet
- Net Off 2004 EpilepsyDocument9 pagesNet Off 2004 EpilepsypastafarianboyNo ratings yet
- Carson C. Chow and Nancy Kopell - Dynamics of Spiking Neurons With Electrical CouplingDocument28 pagesCarson C. Chow and Nancy Kopell - Dynamics of Spiking Neurons With Electrical CouplingJmasnNo ratings yet
- Neuromorphic Photonics: Introduction To Spiking and ExcitabilityDocument5 pagesNeuromorphic Photonics: Introduction To Spiking and Excitabilityrama chaudharyNo ratings yet
- Rox in 2011 RoleDocument15 pagesRox in 2011 RolepastafarianboyNo ratings yet
- Brain Connectivity-A TutorialDocument86 pagesBrain Connectivity-A TutorialcericarNo ratings yet
- Vanrossum2002fast PDFDocument11 pagesVanrossum2002fast PDFpastafarianboyNo ratings yet
- Paul C. Bressloff - Resonantlike Synchronization and Bursting in A Model of Pulse-Coupled Neurons With Active DendritesDocument13 pagesPaul C. Bressloff - Resonantlike Synchronization and Bursting in A Model of Pulse-Coupled Neurons With Active DendritesNeerFamNo ratings yet
- EEG and Brain Connectivity: A TutorialDocument90 pagesEEG and Brain Connectivity: A Tutorialoscar jesu peñaNo ratings yet
- A Modified Cable Formalism For Modeling Neuronal Membranes at High FrequenciesDocument25 pagesA Modified Cable Formalism For Modeling Neuronal Membranes at High Frequenciesdzh12345No ratings yet
- Rate Models For Conductance-Based Cortical Neuronal NetworksDocument33 pagesRate Models For Conductance-Based Cortical Neuronal NetworksFrontiersNo ratings yet
- Dynamics of The Fitzhugh-Nagumo Neuron Model PDFDocument24 pagesDynamics of The Fitzhugh-Nagumo Neuron Model PDFANAMIKA DASNo ratings yet
- KGG Labyrinth Final EPJD 58 181 2010 960kDocument6 pagesKGG Labyrinth Final EPJD 58 181 2010 960kcesar abraham torrico chavezNo ratings yet
- Hansel 1995 SynchronyDocument31 pagesHansel 1995 SynchronypastafarianboyNo ratings yet
- Podlaski Machens 2024Document55 pagesPodlaski Machens 2024ckmachensNo ratings yet
- Hansel 1992 SynchronizationDocument4 pagesHansel 1992 SynchronizationpastafarianboyNo ratings yet
- EEG and BrainDocument110 pagesEEG and BrainGeo MeNo ratings yet
- Neural Networks and Physical Systems With Emergent Collective Computational Abilities HOPFIELD 82Document6 pagesNeural Networks and Physical Systems With Emergent Collective Computational Abilities HOPFIELD 82Kita de AzevedoNo ratings yet
- TMP 958Document12 pagesTMP 958FrontiersNo ratings yet
- Letter: Quasiparticle Engineering and Entanglement Propagation in A Quantum Many-Body SystemDocument11 pagesLetter: Quasiparticle Engineering and Entanglement Propagation in A Quantum Many-Body SystemTuring DanielaNo ratings yet
- The Biomedical Engineering Handbook: Second EditionDocument14 pagesThe Biomedical Engineering Handbook: Second Editionjorgeluis.unknownman667No ratings yet
- Smni83 Dynamics PDFDocument32 pagesSmni83 Dynamics PDFLester IngberNo ratings yet
- Esensor SNR Heter v3Document19 pagesEsensor SNR Heter v3Saul Diaz Infante VelascoNo ratings yet
- 1 s2.0 S0378437104006363 MainDocument9 pages1 s2.0 S0378437104006363 Mainab1699No ratings yet
- Reconfigurable Acceleration of Neural Models With Gap JunctionsDocument4 pagesReconfigurable Acceleration of Neural Models With Gap JunctionsEngr Hafeez QaziNo ratings yet
- Macke 2009 GeneratingDocument28 pagesMacke 2009 GeneratingpastafarianboyNo ratings yet
- Neurons With Graded Response Have Collective Computational Properties Like Those of Two-State NeuronsDocument6 pagesNeurons With Graded Response Have Collective Computational Properties Like Those of Two-State NeuronskivanaNo ratings yet
- Kawabata 1981Document7 pagesKawabata 1981DANIEL ARTEAGA MENDOZANo ratings yet
- Nature and Scope of AI Techniques: Seminar Report Nov2011Document23 pagesNature and Scope of AI Techniques: Seminar Report Nov2011BaneeIshaqueKNo ratings yet
- Static Security Assessment of Power System Using Kohonen Neural Network A. El-SharkawiDocument5 pagesStatic Security Assessment of Power System Using Kohonen Neural Network A. El-Sharkawirokhgireh_hojjatNo ratings yet
- Dynamical Aspects of Kinouchi-Copelli Model: Emergence of Avalanches at CriticalityDocument4 pagesDynamical Aspects of Kinouchi-Copelli Model: Emergence of Avalanches at CriticalityEdward YraitaNo ratings yet
- Efthymios Kallos Et Al - A Multibunch Plasma Wakefield AcceleratorDocument3 pagesEfthymios Kallos Et Al - A Multibunch Plasma Wakefield AcceleratorVasmazxNo ratings yet
- Iliuk, I., Balthazar, J.M., Tusset, A.M., Piqueira, J.R.C., de Pontes, B.R., Felix, J.L.P., Bueno, A.MDocument1 pageIliuk, I., Balthazar, J.M., Tusset, A.M., Piqueira, J.R.C., de Pontes, B.R., Felix, J.L.P., Bueno, A.Mitamar15No ratings yet
- PRE 70 066626 NonlinearSupratransmissionFPUDocument7 pagesPRE 70 066626 NonlinearSupratransmissionFPUJeanette AmayaNo ratings yet
- PhysRevLett 98 104101Document4 pagesPhysRevLett 98 104101cesar abraham torrico chavezNo ratings yet
- Entire Spectrum of Fully Many-Body Localized Systems Using Tensor NetworksDocument12 pagesEntire Spectrum of Fully Many-Body Localized Systems Using Tensor NetworksAnonymous lqvgUsVckNo ratings yet
- Influence of Macrocolumnar EEG On Ca Waves: Lester IngberDocument4 pagesInfluence of Macrocolumnar EEG On Ca Waves: Lester IngberLester IngberNo ratings yet
- Exp 6Document13 pagesExp 6GlenNo ratings yet
- Disentanglement of Local Field Potential Sources by Independent Component AnalysisDocument13 pagesDisentanglement of Local Field Potential Sources by Independent Component AnalysisFrontiersNo ratings yet
- TMP 803 FDocument5 pagesTMP 803 FFrontiersNo ratings yet
- Wave Speed in Excitable Random Networks With Spatially Constrained ConnectionsDocument10 pagesWave Speed in Excitable Random Networks With Spatially Constrained ConnectionsNikita VladimirovNo ratings yet
- S. Coombes and P.C. Bressloff - Solitary Waves in A Model of Dendritic Cable With Active SpinesDocument22 pagesS. Coombes and P.C. Bressloff - Solitary Waves in A Model of Dendritic Cable With Active SpinesNeerFamNo ratings yet
- Vai 1999Document11 pagesVai 1999dsa sdsddsddsNo ratings yet
- (2016) Analysis of Neuronal Spike Trains DeconstructedDocument39 pages(2016) Analysis of Neuronal Spike Trains DeconstructedDiego Soto ChavezNo ratings yet
- Cellular Neural NetworkDocument7 pagesCellular Neural NetworkxasohafNo ratings yet
- The Structure of Multi-Neuron Firing Patterns in Primate RetinaDocument13 pagesThe Structure of Multi-Neuron Firing Patterns in Primate RetinaSebastián GallegosNo ratings yet
- Bayes Optimal Template Matching For Spike Sorting - Combining Fisher Discriminant Analysis With Optimal FilteringDocument21 pagesBayes Optimal Template Matching For Spike Sorting - Combining Fisher Discriminant Analysis With Optimal FilteringatewogboNo ratings yet
- PCMO RRAM For Integrate-and-Fire Neuron in Spiking Neural NetworksDocument4 pagesPCMO RRAM For Integrate-and-Fire Neuron in Spiking Neural NetworksRakesh KumarNo ratings yet
- Anfis NoteDocument15 pagesAnfis NoteNimoNo ratings yet
- Electroencephalographic Field Influence On Calcium Momentum WavesDocument16 pagesElectroencephalographic Field Influence On Calcium Momentum WavesLester IngberNo ratings yet
- Lester Ingber Research, P.O. Box 857, Mclean, Virginia 22101Document11 pagesLester Ingber Research, P.O. Box 857, Mclean, Virginia 22101Lester IngberNo ratings yet
- Segmentation and Edge Detection Based On Spiking Neural Network ModelDocument16 pagesSegmentation and Edge Detection Based On Spiking Neural Network ModelLàm Màu ChannelNo ratings yet
- RBF BP JieeecDocument5 pagesRBF BP JieeecEyad A. FeilatNo ratings yet
- Marder-Goaillard, 2006 Nat Nsci, Homeostasis in Neuron and NetworksDocument12 pagesMarder-Goaillard, 2006 Nat Nsci, Homeostasis in Neuron and NetworksAida OrtegaNo ratings yet
- Electromagnetic Support Arrangement With Threedimensional Control. Ii. ExperimentalDocument3 pagesElectromagnetic Support Arrangement With Threedimensional Control. Ii. Experimentalandres sanchezNo ratings yet
- Jean Faber and Gilson A. Giraldi - Quantum Models For Artifcial Neural NetworkDocument8 pagesJean Faber and Gilson A. Giraldi - Quantum Models For Artifcial Neural Networkdcsi3No ratings yet
- Letters: Evidence For Wavelike Energy Transfer Through Quantum Coherence in Photosynthetic SystemsDocument5 pagesLetters: Evidence For Wavelike Energy Transfer Through Quantum Coherence in Photosynthetic SystemsJames LimNo ratings yet
- PhysRevX 9 031009Document21 pagesPhysRevX 9 031009Marc de MiguelNo ratings yet
- Negative Mass and Negative Refractive Index in Atom Nuclei - Nuclear Wave Equation - Gravitational and Inertial Control: Part 5: Gravitational and Inertial Control, #5From EverandNegative Mass and Negative Refractive Index in Atom Nuclei - Nuclear Wave Equation - Gravitational and Inertial Control: Part 5: Gravitational and Inertial Control, #5No ratings yet
- Negative Mass and Negative Refractive Index in Atom Nuclei - Nuclear Wave Equation - Gravitational and Inertial Control: Part 2: Gravitational and Inertial Control, #2From EverandNegative Mass and Negative Refractive Index in Atom Nuclei - Nuclear Wave Equation - Gravitational and Inertial Control: Part 2: Gravitational and Inertial Control, #2No ratings yet
- Electromgnetism Homework 5 Edward Iraita SDocument24 pagesElectromgnetism Homework 5 Edward Iraita SEdward YraitaNo ratings yet
- Electromgnetism Homework 5 Edward Iraita SDocument24 pagesElectromgnetism Homework 5 Edward Iraita SEdward YraitaNo ratings yet
- Heald - Indb 1 Heald - Indb 1 1/30/09 2:12:22 PM 1/30/09 2:12:22 PMDocument149 pagesHeald - Indb 1 Heald - Indb 1 1/30/09 2:12:22 PM 1/30/09 2:12:22 PMEdward YraitaNo ratings yet
- Electromagnetism Homework 2 Edward Iraita SDocument22 pagesElectromagnetism Homework 2 Edward Iraita SEdward YraitaNo ratings yet
- Dynamical Aspects of Kinouchi-Copelli Model: Emergence of Avalanches at CriticalityDocument4 pagesDynamical Aspects of Kinouchi-Copelli Model: Emergence of Avalanches at CriticalityEdward YraitaNo ratings yet
- 03 Randomgraphs PDFDocument66 pages03 Randomgraphs PDFEdward YraitaNo ratings yet
- Lorentz Attractor Report Ben ReedDocument16 pagesLorentz Attractor Report Ben ReedEdward YraitaNo ratings yet
- Towards A Security Architecture For SubstationsDocument6 pagesTowards A Security Architecture For SubstationsSofyan AndikaNo ratings yet
- 2301B Dental Unit Manual-BeingDocument22 pages2301B Dental Unit Manual-BeingMr.SDNo ratings yet
- Chapter 13: Spreadsheets: Activity 1Document4 pagesChapter 13: Spreadsheets: Activity 1Suzanna OllivierreNo ratings yet
- Hooke's LawDocument19 pagesHooke's LawSary LimNo ratings yet
- To Find The Refractive Index of WaterDocument13 pagesTo Find The Refractive Index of WaterUtkarshNo ratings yet
- Templates in CPPDocument4 pagesTemplates in CPPElango PerumalNo ratings yet
- Solr PPTDocument30 pagesSolr PPTAlok KumarNo ratings yet
- Pumps and Pumping SystemsDocument88 pagesPumps and Pumping SystemssrathinakumarNo ratings yet
- Simplified English For Aircraft Workcards: Human Factors and Ergonomics Society Annual Meeting Proceedings October 1996Document17 pagesSimplified English For Aircraft Workcards: Human Factors and Ergonomics Society Annual Meeting Proceedings October 1996MariamVTNo ratings yet
- Interview Questions On ESXi 5.5Document1 pageInterview Questions On ESXi 5.5jay_tiwariSITNo ratings yet
- Approved Final Specification Lan ExtenderDocument13 pagesApproved Final Specification Lan ExtenderShubham KumarNo ratings yet
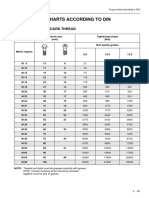
- 2.3 Torque Charts According To Din: 2.3.1 Metric Standard ThreadDocument3 pages2.3 Torque Charts According To Din: 2.3.1 Metric Standard Threadamplefex medinaNo ratings yet
- DLL - Mathematics 6 - Q1 - W9Document4 pagesDLL - Mathematics 6 - Q1 - W9Elc Elc ElcNo ratings yet
- Wood Element Axial Bending EC5Document2 pagesWood Element Axial Bending EC5nicolaemariusNo ratings yet
- Taekwondo : Testing SyllabusDocument18 pagesTaekwondo : Testing SyllabusJulio OlmedoNo ratings yet
- J Station ManualDocument39 pagesJ Station ManualRas Bongo Natty TafariNo ratings yet
- Intermediate MicroeconomicsDocument36 pagesIntermediate MicroeconomicsKintu GeraldNo ratings yet
- Transformation - 2DDocument93 pagesTransformation - 2DKashika MehtaNo ratings yet
- FINALE BlaaaaaDocument55 pagesFINALE BlaaaaaYvanah CapuyanNo ratings yet
- Common Network Cable: Learning Activity SheetDocument7 pagesCommon Network Cable: Learning Activity SheetRamlede BenosaNo ratings yet
- FlowconME Valves Catalogue 13Document1 pageFlowconME Valves Catalogue 13mohansaf0% (1)
- Power Transformer MaintenanceDocument19 pagesPower Transformer Maintenanceadnan83% (6)
- B Tech Mechanical Engineering Syllabus For Batch 2013-14Document30 pagesB Tech Mechanical Engineering Syllabus For Batch 2013-14ra44993541No ratings yet
- Sulsim Script FinalDocument10 pagesSulsim Script FinalMuhammad NaeemNo ratings yet
- Enzyme: Ayesha Shafi Pharm-D, (P.U.), M. Phil. Pharmaceutical Chemistry (P.U.)Document34 pagesEnzyme: Ayesha Shafi Pharm-D, (P.U.), M. Phil. Pharmaceutical Chemistry (P.U.)Shafaqat Ghani Shafaqat Ghani100% (2)
- ChronoamperometryDocument7 pagesChronoamperometrybettypaz89100% (1)
- Geometric مراجعة جزء الـDocument2 pagesGeometric مراجعة جزء الـamin alzuraikiNo ratings yet
- EMI6000 Cal 6Document2 pagesEMI6000 Cal 6John DolanNo ratings yet