You might also like
- Starving Cancer Cells: Evidence-Based Strategies to Slow Cancer Progression: A Selection of Readings for Health Services ProvidersFrom EverandStarving Cancer Cells: Evidence-Based Strategies to Slow Cancer Progression: A Selection of Readings for Health Services ProvidersNo ratings yet
- Wajib BacaDocument17 pagesWajib BacaRum Afida RasfaNo ratings yet
- Jurnal Tumor Metastasis Paru Hepar Dari PancreasDocument15 pagesJurnal Tumor Metastasis Paru Hepar Dari PancreaslucyishakNo ratings yet
- Glucose Metabolism in Cancer and The Role of Ketogenic DietDocument9 pagesGlucose Metabolism in Cancer and The Role of Ketogenic DietЛилия ПоляковаNo ratings yet
- SasdsadsaDocument11 pagesSasdsadsaJorge MariaNo ratings yet
- The Evolving Role of The Endocannabinoid System in Gynaecological CancerDocument19 pagesThe Evolving Role of The Endocannabinoid System in Gynaecological CancernusantaramedikasolusindoNo ratings yet
- Cancers: Translational Research in Ovarian CancerDocument2 pagesCancers: Translational Research in Ovarian CancerRomina DatuNo ratings yet
- 10.1038@s41575 018 0069 7Document16 pages10.1038@s41575 018 0069 7Ronald Echeverria IbazetaNo ratings yet
- 2023 Monteleone Tagerted therapy of IL34 as a prmosing approach to overcome cancer resistanceDocument12 pages2023 Monteleone Tagerted therapy of IL34 as a prmosing approach to overcome cancer resistancephilippe.lacroix38490No ratings yet
- El Cáncer en La Función MitocondrialDocument4 pagesEl Cáncer en La Función MitocondrialJhan SaavedraNo ratings yet
- Ol 18 1 487 PDFDocument12 pagesOl 18 1 487 PDFCasiana GuiNo ratings yet
- NIH Public Access: Author ManuscriptDocument17 pagesNIH Public Access: Author ManuscriptmarcussiNo ratings yet
- Thyroid Cancer: Burden of Illness and Management of Disease: J o U R N A L o F C A N C e RDocument7 pagesThyroid Cancer: Burden of Illness and Management of Disease: J o U R N A L o F C A N C e RRum Afida RasfaNo ratings yet
- The Progress of Methylation Regulation in Gene ExpDocument12 pagesThe Progress of Methylation Regulation in Gene ExpDavidNo ratings yet
- Pancreatic CancerDocument13 pagesPancreatic CancerFA MonterNo ratings yet
- Bergh 2013Document4 pagesBergh 2013bixagif369No ratings yet
- BPJ Vol 14 No 4 P 1993-2005Document13 pagesBPJ Vol 14 No 4 P 1993-2005Dr. Shraddha YadavNo ratings yet
- Metabolomics in Oncology: Gurparsad Singh Suri - Gurleen Kaur - Giuseppina M. Carbone - Dheeraj ShindeDocument13 pagesMetabolomics in Oncology: Gurparsad Singh Suri - Gurleen Kaur - Giuseppina M. Carbone - Dheeraj ShindePreethi SridharanNo ratings yet
- Apicella 2015Document16 pagesApicella 2015bawoji1763No ratings yet
- Cancer Anaerobic GlycolysisDocument16 pagesCancer Anaerobic GlycolysisMilaNo ratings yet
- Endometrialcancer: Obesity, Genetics, and Targeted AgentsDocument17 pagesEndometrialcancer: Obesity, Genetics, and Targeted AgentsEftychia GkikaNo ratings yet
- Cancers 15 03942Document23 pagesCancers 15 03942Monique XyztNo ratings yet
- Genetic-Guidedrisk Assessmentand Managementofthyroidcancer: Mingzhao XingDocument16 pagesGenetic-Guidedrisk Assessmentand Managementofthyroidcancer: Mingzhao XingPedro Gómez RNo ratings yet
- Review 2015Document9 pagesReview 2015akhotnik.tndNo ratings yet
- Cancers 02 02084Document14 pagesCancers 02 02084MAGNo ratings yet
- Idrd 30 2144541Document18 pagesIdrd 30 2144541Uzma KhanNo ratings yet
- Thyroid Carcinoma: Phenotypic Features, Underlying Biology and Potential Relevance For Targeting TherapyDocument25 pagesThyroid Carcinoma: Phenotypic Features, Underlying Biology and Potential Relevance For Targeting TherapyrakaNo ratings yet
- W de WenisimoDocument18 pagesW de WenisimoBJ CarminatorNo ratings yet
- The Metabolic Interplay between Cancer and Other Diseases ReviewDocument13 pagesThe Metabolic Interplay between Cancer and Other Diseases ReviewPancho ArancibiaNo ratings yet
- 1 s2.0 S1044579X14000510 MainDocument9 pages1 s2.0 S1044579X14000510 MainGwzthabvo JaimeNo ratings yet
- Role & Future of SBRT in OMDDocument14 pagesRole & Future of SBRT in OMDMayur MayankNo ratings yet
- Propofol-TIVA Versus Inhalational Anesthesia For Cancer SurgeryDocument9 pagesPropofol-TIVA Versus Inhalational Anesthesia For Cancer SurgerygonjdelzNo ratings yet
- PART 2 Coran Pediatric Surgery, 7th Ed PDFDocument350 pagesPART 2 Coran Pediatric Surgery, 7th Ed PDFRamona Cerasela Petreaca100% (4)
- Molecular Pathogenesis and Mechanisms of Thyroid Cancer: Mingzhao XingDocument16 pagesMolecular Pathogenesis and Mechanisms of Thyroid Cancer: Mingzhao XingTaeyomiNo ratings yet
- 10 1016@j Canlet 2020 10 029Document54 pages10 1016@j Canlet 2020 10 029Graziane PortelaNo ratings yet
- A Literature Review of Complementary and Alternative Medicine Use by Colorectal Cancer PatientsDocument4 pagesA Literature Review of Complementary and Alternative Medicine Use by Colorectal Cancer Patientsgw0a869xNo ratings yet
- Epithelial To Mesenchymal Transition and Cell Biology of Molecular Regulation in Endometrial CarcinogenesisDocument17 pagesEpithelial To Mesenchymal Transition and Cell Biology of Molecular Regulation in Endometrial CarcinogenesisMelati HasnailNo ratings yet
- GatenbyGillies2007 MicroenvmodelDocument6 pagesGatenbyGillies2007 Microenvmodelpanna1No ratings yet
- Metabolismo Da Glicose Na Plasticidade, Diagnóstico e Tratamento Do TumorDocument10 pagesMetabolismo Da Glicose Na Plasticidade, Diagnóstico e Tratamento Do TumorvalquiriapolNo ratings yet
- Cancer-Associated Fibroblasts in Breast Cancer ChallengesDocument34 pagesCancer-Associated Fibroblasts in Breast Cancer ChallengesANA PAULANo ratings yet
- The Cell Biological Basis of Cancer - ThesisDocument50 pagesThe Cell Biological Basis of Cancer - ThesisRamesh MahakurNo ratings yet
- Higher Rate of BRAF Mutation in Papillary Thyroid Cancer Over TimeDocument6 pagesHigher Rate of BRAF Mutation in Papillary Thyroid Cancer Over TimeEnda RafiqohNo ratings yet
- Selle2015 PDFDocument12 pagesSelle2015 PDFatikha apriliaNo ratings yet
- Cancer 55Document9 pagesCancer 55Bilal SaeedNo ratings yet
- Metabolic Signatures of Human Breast CancerDocument11 pagesMetabolic Signatures of Human Breast CancerMehedi HossainNo ratings yet
- MedComm - 2020 - FuDocument33 pagesMedComm - 2020 - Fuasfs92323No ratings yet
- Bussines EngineringDocument9 pagesBussines EngineringWhulan Rhana TonapaNo ratings yet
- Sample Research Paper On Colon CancerDocument6 pagesSample Research Paper On Colon Cancerihprzlbkf100% (1)
- The Role of CD44 in Cancer Chemoresistance A Concise ReviewDocument13 pagesThe Role of CD44 in Cancer Chemoresistance A Concise ReviewZihao WuNo ratings yet
- Journal of Theoretical Biology: Gibin G. Powathil, Kirsty E. Gordon, Lydia A. Hill, Mark A.J. ChaplainDocument19 pagesJournal of Theoretical Biology: Gibin G. Powathil, Kirsty E. Gordon, Lydia A. Hill, Mark A.J. ChaplainLal RintluangaNo ratings yet
- 2017 Article 52-2Document9 pages2017 Article 52-2Lorena RamosNo ratings yet
- Urea Cycle Sustains Cellular Energetics Upon EGFR Inhibition in EGFR Mutant NSCLCDocument36 pagesUrea Cycle Sustains Cellular Energetics Upon EGFR Inhibition in EGFR Mutant NSCLCJaja LazaroNo ratings yet
- Bone Metastases From Thyroid Carcinoma of Follicular Origin A Single Institutional ExperienceDocument6 pagesBone Metastases From Thyroid Carcinoma of Follicular Origin A Single Institutional ExperienceChavdarNo ratings yet
- AJHO Apr15 10 ThyroidDocument5 pagesAJHO Apr15 10 ThyroidAura RachmawatiNo ratings yet
- Molecular Profiling in Muscle-Invasive Bladder Cancer: More Than The Sum of Its PartsDocument11 pagesMolecular Profiling in Muscle-Invasive Bladder Cancer: More Than The Sum of Its PartsJoel ChávezdrumsNo ratings yet
- 41 All inDocument11 pages41 All inbawoji1763No ratings yet
- Genetics May Guide Gastric Cancer TreatmentDocument26 pagesGenetics May Guide Gastric Cancer Treatmentjomigp01No ratings yet
- PL 00011733Document9 pagesPL 00011733Monica CabaNo ratings yet
- The Cancer HandbookDocument1,529 pagesThe Cancer HandbookAsimaJulianaSiregarNo ratings yet
- Emerging Targeted Agents in Endometrial Cancer Treatment: Critical ReviewDocument4 pagesEmerging Targeted Agents in Endometrial Cancer Treatment: Critical Reviewmikk85No ratings yet
- Hyper Pro Lac Tine MiaDocument47 pagesHyper Pro Lac Tine Miakharbosham100% (3)
- Menu: Print ShareDocument37 pagesMenu: Print ShareAndreea CimpoiNo ratings yet
- Urn - Isbn - 978 952 61 2713 2Document136 pagesUrn - Isbn - 978 952 61 2713 2dewi najiraNo ratings yet
- Akp Important Guidelines 22 Guidlines Brief FinalDocument55 pagesAkp Important Guidelines 22 Guidlines Brief FinalRaheelNo ratings yet
- Guillain - Barre - SyndromeDocument18 pagesGuillain - Barre - SyndromeRAMNo ratings yet
- Neuropathic PainDocument40 pagesNeuropathic PainHasmirahNo ratings yet
- Hyperthermia HandoutDocument1 pageHyperthermia Handoutapi-332422697No ratings yet
- Rabies PresentationDocument23 pagesRabies PresentationAvdhesh YadavNo ratings yet
- 18-RECK MOTOMED Movement Therapy Viva 2 - CompressPdfDocument8 pages18-RECK MOTOMED Movement Therapy Viva 2 - CompressPdfFardiyan SyafriNo ratings yet
- Migraine DiaryDocument6 pagesMigraine DiaryhumphryNo ratings yet
- Rectal Suppository Procedure With Rationale 1Document5 pagesRectal Suppository Procedure With Rationale 1yuuki konnoNo ratings yet
- Remedy Responses (LMS)Document1 pageRemedy Responses (LMS)Roger Knight100% (3)
- Cancer InnovationsDocument60 pagesCancer Innovationseustaquiocajaenpena100% (2)
- Eastern Health 30 Minute 4 Hour Rule 2017 PDFDocument1 pageEastern Health 30 Minute 4 Hour Rule 2017 PDFRosel InaNo ratings yet
- Male Pelvic Floor - Advanced Massage and BodyworkDocument6 pagesMale Pelvic Floor - Advanced Massage and Bodyworkpujarze2No ratings yet
- Approach To Polyarthritis For The Primary Care Physician: Arielle Freilich, DO, PGY2 & Helaine Larsen, DODocument8 pagesApproach To Polyarthritis For The Primary Care Physician: Arielle Freilich, DO, PGY2 & Helaine Larsen, DOLydia IsaacNo ratings yet
- Contact Tracing Form San LuisDocument3 pagesContact Tracing Form San LuisPoblacion 04 San LuisNo ratings yet
- He Effectiveness of Brandt Daroff Exercises On Vertigo Events in Subjects of Vertigo PatientsDocument4 pagesHe Effectiveness of Brandt Daroff Exercises On Vertigo Events in Subjects of Vertigo PatientsRadhikaNo ratings yet
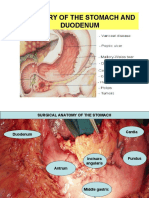
- Surgery of The Stomach and DuodenumDocument60 pagesSurgery of The Stomach and DuodenumTarikNo ratings yet
- Hannah SchizopreniaDocument5 pagesHannah SchizopreniaEliezah RodriguezNo ratings yet
- ALS Brochure 508compDocument24 pagesALS Brochure 508compidaayupranitaNo ratings yet
- Dental Management of Patients Under Anticoagulant TherapyDocument15 pagesDental Management of Patients Under Anticoagulant TherapyThaer ZabenNo ratings yet
- CNS Version 2.0Document127 pagesCNS Version 2.0Neeladri DawnNo ratings yet
- Sideroblastic Anemia Guide: Causes, Symptoms & TypesDocument18 pagesSideroblastic Anemia Guide: Causes, Symptoms & TypesSonam Joshi100% (2)
- Fluid Balance Nursing: Assess, Chart, Prevent ImbalanceDocument11 pagesFluid Balance Nursing: Assess, Chart, Prevent ImbalanceEla GabrielaNo ratings yet
- Alltest CatalogDocument22 pagesAlltest CatalogAngelo PlataNo ratings yet
- PrioritizationDocument9 pagesPrioritizationmiss RN100% (4)
- Abdominal TraumaDocument52 pagesAbdominal TraumaDONALD UNASHENo ratings yet
- Scleroderma: Quick Recertific Ation SeriesDocument2 pagesScleroderma: Quick Recertific Ation SeriesvieeveeNo ratings yet
- Snoreben - Anti Snoring DeviceDocument14 pagesSnoreben - Anti Snoring DevicerepharmhealthcareNo ratings yet