You might also like
- Polli Et Al, 2009, An Unexpected Synergist Role of P-Glycoprotein and Breast Cancer Resistance Protein On The CentralDocument4 pagesPolli Et Al, 2009, An Unexpected Synergist Role of P-Glycoprotein and Breast Cancer Resistance Protein On The Centraldhania novitasariNo ratings yet
- The Pharmacodynamics and Pharmacokinetics of Mivacurium in ChildrenDocument7 pagesThe Pharmacodynamics and Pharmacokinetics of Mivacurium in ChildrenluyawinNo ratings yet
- Zhou 2007Document11 pagesZhou 2007Rais RyuzakiNo ratings yet
- Pluripotent Anti-Inflammatory Immunomodulatory Effects of Papaverine Against Cerebral Ischemic-Reperfusion InjuryDocument23 pagesPluripotent Anti-Inflammatory Immunomodulatory Effects of Papaverine Against Cerebral Ischemic-Reperfusion InjuryDragomir MirunaNo ratings yet
- Cheng Lu., Et Al. 2016Document7 pagesCheng Lu., Et Al. 2016Ubaida AssalwaNo ratings yet
- Múltiple cerebrointraventricular administrationDocument5 pagesMúltiple cerebrointraventricular administrationivan941919No ratings yet
- Article 4Document13 pagesArticle 4ירדן לויןNo ratings yet
- 28sici 291097 4547 2819961101 2946 3A3 3C316 3A 3aaid jnr4 3e3.0.co 3B2 NDocument8 pages28sici 291097 4547 2819961101 2946 3A3 3C316 3A 3aaid jnr4 3e3.0.co 3B2 NLata DeshmukhNo ratings yet
- Journal AphasiaDocument9 pagesJournal AphasiaChintya Anugrah SuhendraNo ratings yet
- J Pharmacol Exp Ther 2003 Zhao 246 53Document8 pagesJ Pharmacol Exp Ther 2003 Zhao 246 53mcgilicuttyNo ratings yet
- Epilepsy ResearchDocument9 pagesEpilepsy ResearchDiego Carmona MontielNo ratings yet
- James M. Radke Et Al - Atypical Antipsychotic Drugs Selectively Increase Neurotensin Efflux in Dopamine Terminal RegionsDocument3 pagesJames M. Radke Et Al - Atypical Antipsychotic Drugs Selectively Increase Neurotensin Efflux in Dopamine Terminal RegionsLonkesNo ratings yet
- Drug Metab Dispos 2005 Maher 956 62Document7 pagesDrug Metab Dispos 2005 Maher 956 62Anshari StressNo ratings yet
- TCI Propofol ClearanceDocument9 pagesTCI Propofol ClearanceJoanna GlezNo ratings yet
- Desferrioxamine Induces Delayed Tolerance Against Cerebral Ischemia in Vivo and in VitroDocument6 pagesDesferrioxamine Induces Delayed Tolerance Against Cerebral Ischemia in Vivo and in VitroJephte LaputNo ratings yet
- 2013 AapsjDocument8 pages2013 AapsjSCYoungNo ratings yet
- ACTH Treatment For Management of Nephrotic SyndromeDocument16 pagesACTH Treatment For Management of Nephrotic SyndromeCalvin Tanuwijaya Stick BolaNo ratings yet
- Immunohistochemically Detected Expression of 3 Major Genes (CDKN2A/p16, TP53, and SMAD4/DPC4) Strongly Predicts Survival in Patients With Resectable Pancreatic CancerDocument11 pagesImmunohistochemically Detected Expression of 3 Major Genes (CDKN2A/p16, TP53, and SMAD4/DPC4) Strongly Predicts Survival in Patients With Resectable Pancreatic CancerSisca Dwi AgustinaNo ratings yet
- Pier Vincenzo Piazza Et Al - Glucocorticoids Have State-Dependent Stimulant Effects On The Mesencephalic Dopaminergic TransmissionDocument5 pagesPier Vincenzo Piazza Et Al - Glucocorticoids Have State-Dependent Stimulant Effects On The Mesencephalic Dopaminergic TransmissionSour60No ratings yet
- ParkinsonDocument9 pagesParkinsonAnonymous lwYEylQhNo ratings yet
- Debabrata JTM 2013Document13 pagesDebabrata JTM 2013Tarak Nath KhatuaNo ratings yet
- 1 s2.0 S0753332221010702 MainDocument18 pages1 s2.0 S0753332221010702 MainGopalaKrishnanNo ratings yet
- Poster Viewing I Tuesday 1 May 2018 Iii75Document2 pagesPoster Viewing I Tuesday 1 May 2018 Iii75Firdaus Septhy ArdhyanNo ratings yet
- Systematic Search For Variation in The Human Norepinephrine Transporter GeneDocument10 pagesSystematic Search For Variation in The Human Norepinephrine Transporter GeneAltaicaNo ratings yet
- 402 FullDocument7 pages402 Fullyohanes0gadiNo ratings yet
- WinsaamDocument11 pagesWinsaamdinisaaruniNo ratings yet
- Interictal, Circulating Sphingolipids in Women With Episodic MigraineDocument11 pagesInterictal, Circulating Sphingolipids in Women With Episodic MigraineAnonymous bx7YQvNo ratings yet
- Global REACH 2018: Volume Regulation in High-Altitude Andeans With and Without Chronic Mountain SicknessDocument9 pagesGlobal REACH 2018: Volume Regulation in High-Altitude Andeans With and Without Chronic Mountain SicknessFiorella LeyvaNo ratings yet
- J Pharmacol Exp Ther 1999 Hesselink 543 50Document8 pagesJ Pharmacol Exp Ther 1999 Hesselink 543 50CLPHtheoryNo ratings yet
- BPH 174 3727Document22 pagesBPH 174 3727taniasolisNo ratings yet
- Macroprolactinemia Revisited A Study On 106 Patients JcemDocument8 pagesMacroprolactinemia Revisited A Study On 106 Patients JcemPaulo Eduardo CampanaNo ratings yet
- Espinoza Et Al 2014Document9 pagesEspinoza Et Al 2014niltonNo ratings yet
- PIIS00150282990060Document4 pagesPIIS00150282990060lilou.magnaboscoNo ratings yet
- Original Article: High Expression of WDR1 in Primary Glioblastoma Is Associated With Poor PrognosisDocument12 pagesOriginal Article: High Expression of WDR1 in Primary Glioblastoma Is Associated With Poor PrognosisALINo ratings yet
- Glial Cell Line-Derived Neurotrophic Factor Protects Against Ischemia-Induced Injury in The Cerebral CortexDocument8 pagesGlial Cell Line-Derived Neurotrophic Factor Protects Against Ischemia-Induced Injury in The Cerebral Cortexranu anggaraNo ratings yet
- Acm 2014 5390 AbstractDocument2 pagesAcm 2014 5390 Abstractharshadpardeshi45No ratings yet
- Pre-Clinical Study of Induced Pluripotent Stem Cell-Derived Dopaminergic Progenitor Cells For Parkinson's DiseaseDocument14 pagesPre-Clinical Study of Induced Pluripotent Stem Cell-Derived Dopaminergic Progenitor Cells For Parkinson's DiseaseMuhammad Hamzah SyahrirNo ratings yet
- Cardioprotective Effects of Curcumin NanocompositeDocument10 pagesCardioprotective Effects of Curcumin NanocompositeAisha MurtazaNo ratings yet
- 1756 0500 6 477 PDFDocument6 pages1756 0500 6 477 PDFanon_118437533No ratings yet
- Alan Breier Et Al - Association of Ketamine-Induced Psychosis With Focal Activation of The Prefrontal Cortex in Healthy VolunteersDocument7 pagesAlan Breier Et Al - Association of Ketamine-Induced Psychosis With Focal Activation of The Prefrontal Cortex in Healthy VolunteersCortate15gNo ratings yet
- Integrated Analysis of Ultra-Deep ProteomesDocument20 pagesIntegrated Analysis of Ultra-Deep ProteomesMAURICIO FLORESNo ratings yet
- Li 2021Document8 pagesLi 2021Intan Adevia RosnaritaNo ratings yet
- Codon-Optimized Human Sodium Iodide Symporter (opt-hNIS) As A Sensitive Reporter and Efficient Therapeutic GeneDocument11 pagesCodon-Optimized Human Sodium Iodide Symporter (opt-hNIS) As A Sensitive Reporter and Efficient Therapeutic GeneAwesoome PocoyoNo ratings yet
- 1 s2.0 S1936523312800428 MainDocument9 pages1 s2.0 S1936523312800428 MainJessica GintingNo ratings yet
- Babalik. Serum drug concentration Turkey. IJTLD Dec 13Document7 pagesBabalik. Serum drug concentration Turkey. IJTLD Dec 13Vigilancia EpidemiologicaNo ratings yet
- Cerebral Dopamine Neurotrophic Factor Regulates Multiple Neuronal Subtypes and BehaviorDocument19 pagesCerebral Dopamine Neurotrophic Factor Regulates Multiple Neuronal Subtypes and BehaviorCONSTANZA TERESA BERNA ARAYANo ratings yet
- Dopamina e GagueiraDocument5 pagesDopamina e GagueiraMaisa Tatiana CasarinNo ratings yet
- Long-Term Clinical Outcome of Fetal Cell Transplantation For Parkinson Disease Two Case ReportsDocument5 pagesLong-Term Clinical Outcome of Fetal Cell Transplantation For Parkinson Disease Two Case Reportsjust for download matterNo ratings yet
- MitochondrialDocument13 pagesMitochondrialAndrea CamachoNo ratings yet
- Ref 47 hasil291096-862829963C2173Aaid-ajmg173B2-0Document5 pagesRef 47 hasil291096-862829963C2173Aaid-ajmg173B2-0Fitri anaNo ratings yet
- Clinical Laboratory Analysis - 2012 - Hashad - Free Circulating Tumor DNA as a Diagnostic Marker for Breast CancerDocument6 pagesClinical Laboratory Analysis - 2012 - Hashad - Free Circulating Tumor DNA as a Diagnostic Marker for Breast Cancerfaraz.mirza1No ratings yet
- In Neonates of 28 Weeks' Gestation Phase 1 Trial of 4 Thyroid Hormone Regimens For Transient HypothyroxinemiaDocument13 pagesIn Neonates of 28 Weeks' Gestation Phase 1 Trial of 4 Thyroid Hormone Regimens For Transient Hypothyroxinemiaapi-28255451No ratings yet
- Identification of Normal Blood Pressure in Different Age GroupDocument8 pagesIdentification of Normal Blood Pressure in Different Age GroupmadhurocksktmNo ratings yet
- 17-Estradiol Attenuates Hippocampal Neuronal LossDocument9 pages17-Estradiol Attenuates Hippocampal Neuronal LossrodrigounitedNo ratings yet
- HronoterapijaDocument12 pagesHronoterapijaSlavicaNo ratings yet
- M. Balíková Et Al - Kinetic Profiles of Incorporation of Some Psychedelics Into Brain Tissue After Subcutaneous Application To RatsDocument6 pagesM. Balíková Et Al - Kinetic Profiles of Incorporation of Some Psychedelics Into Brain Tissue After Subcutaneous Application To RatsSmokeysamNo ratings yet
- 1 s2.0 S0006322397002953nlnl Nnnmain PDFDocument3 pages1 s2.0 S0006322397002953nlnl Nnnmain PDFElisa SalakayNo ratings yet
- The Effect of Polysorbate 80 On Brain Uptake and Analgesic Effect of D-KyotorphinDocument7 pagesThe Effect of Polysorbate 80 On Brain Uptake and Analgesic Effect of D-KyotorphinfrankNo ratings yet
- Hot Spots in Prion Protein For Pathogenic Conversion: SI Fig. 4bDocument6 pagesHot Spots in Prion Protein For Pathogenic Conversion: SI Fig. 4bVishal PatilNo ratings yet
- 2 2 1Document14 pages2 2 1LaurenGoNo ratings yet
- The International Workshop On Meibomian GlandDocument44 pagesThe International Workshop On Meibomian GlandPutri kartiniNo ratings yet
- BARANGAY MONTHLY INVENTORY OF VACCINATED POPULATIONDocument108 pagesBARANGAY MONTHLY INVENTORY OF VACCINATED POPULATIONAna Marie BergoniaNo ratings yet
- Didanosine PDFDocument3 pagesDidanosine PDFRosher Deliman JanoyanNo ratings yet
- Application of Health Belief Model in NursingDocument4 pagesApplication of Health Belief Model in NursingAnusha Verghese100% (1)
- Price Needs Updating, Should We Call The Police?: C M Bleakley, P Glasgow, D C MacauleyDocument2 pagesPrice Needs Updating, Should We Call The Police?: C M Bleakley, P Glasgow, D C MacauleyAna BelchiorNo ratings yet
- (Book 4) Collins Parker, Mia - PCOS Nutrition - A Complete PCOS Diet Book With 4 Week Meal Plan and 4 Week Fitness Exercise Plan To Reduce Weight and Prevent Diabetes. PCOS Causes, Symptoms and HolDocument134 pages(Book 4) Collins Parker, Mia - PCOS Nutrition - A Complete PCOS Diet Book With 4 Week Meal Plan and 4 Week Fitness Exercise Plan To Reduce Weight and Prevent Diabetes. PCOS Causes, Symptoms and HolAndreea Daniela100% (4)
- Health6 - q1 - Mod5 - Identifies Community Health Resources and Facilities - FINAL.Document24 pagesHealth6 - q1 - Mod5 - Identifies Community Health Resources and Facilities - FINAL.vamps sier100% (1)
- WHO Smallpox Eradication Programme Strategy EvolutionDocument56 pagesWHO Smallpox Eradication Programme Strategy EvolutionRahul NetragaonkarNo ratings yet
- Decreased Cardiac OutputDocument9 pagesDecreased Cardiac OutputRae AnnNo ratings yet
- MorsDocument13 pagesMorsBSRT1A BERBANO, IAN JEWEL M.No ratings yet
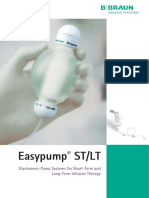
- Easypump ST/LT: Elastomeric Pump Systems For Short-Term and Long-Term Infusion TherapyDocument2 pagesEasypump ST/LT: Elastomeric Pump Systems For Short-Term and Long-Term Infusion TherapyTedyJaswadiNo ratings yet
- ESSENTIAL NEWBORN CARE CHECKLIST UpdatedDocument2 pagesESSENTIAL NEWBORN CARE CHECKLIST UpdatedKristil Chavez100% (2)
- Doctors - Dermatologists Area Phone: Tuesday, 13 November 2007 13:51Document18 pagesDoctors - Dermatologists Area Phone: Tuesday, 13 November 2007 13:51pinakindpatelNo ratings yet
- Old Canalicular Laceration Repair A Retrospective Study ofDocument6 pagesOld Canalicular Laceration Repair A Retrospective Study ofMuh Syarifullah ANo ratings yet
- Ectopic PregnancyDocument25 pagesEctopic Pregnancyfred_chandh88No ratings yet
- ComaDocument13 pagesComaMamoon J. MehdiNo ratings yet
- Kaufmann Protocol guidelines for anti-aging regimensDocument3 pagesKaufmann Protocol guidelines for anti-aging regimenszlautarevic-1No ratings yet
- Family Nursing Care PlanDocument7 pagesFamily Nursing Care PlanRebecca Caga SarmientoNo ratings yet
- Drug StudyDocument7 pagesDrug StudyLA GomezNo ratings yet
- Continuum of Care. Simplified Workflow.: Introducing Trilogy EV300Document3 pagesContinuum of Care. Simplified Workflow.: Introducing Trilogy EV300Shubhobrata BhattacherjeeNo ratings yet
- Neuro-Ophthalmology: Introduction: James Goodwin, MD (Attending)Document4 pagesNeuro-Ophthalmology: Introduction: James Goodwin, MD (Attending)Mariano FioreNo ratings yet
- Cebu Institute of Technology - University: Nursing Care PlanDocument2 pagesCebu Institute of Technology - University: Nursing Care PlanSergiNo ratings yet
- 1 Dental Caries PDFDocument38 pages1 Dental Caries PDFmayang putriNo ratings yet
- British Orthoptic Journal 2002Document9 pagesBritish Orthoptic Journal 2002roelkloosNo ratings yet
- Carcinoma of Prostate: Patho Ogy - Chapter &Document29 pagesCarcinoma of Prostate: Patho Ogy - Chapter &HAMIDNo ratings yet
- Bedside Hemodynamic MonitoringDocument26 pagesBedside Hemodynamic MonitoringBrad F LeeNo ratings yet
- Drug Study DexamethasoneDocument4 pagesDrug Study Dexamethasoneamal abdulrahmanNo ratings yet
- Cystic FibrosisDocument20 pagesCystic FibrosisIndrajit BaruaNo ratings yet
- Fenotipos de BronquiolitisDocument14 pagesFenotipos de BronquiolitisJohann MuñozNo ratings yet