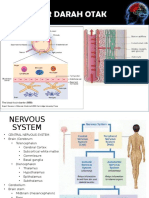
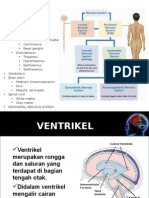
You might also like
- Upper GI Bleeding After Hip and Knee ArthroplastyDocument1 pageUpper GI Bleeding After Hip and Knee ArthroplastyZiad AlazthaNo ratings yet
- Association of Glycemic Status With Bone Turnover Markers in Type 2 Diabetes MellitusDocument5 pagesAssociation of Glycemic Status With Bone Turnover Markers in Type 2 Diabetes MellitusZiad AlazthaNo ratings yet
- 008 NicwodnDocument2 pages008 NicwodnZiad AlazthaNo ratings yet
- Beta His TinDocument1 pageBeta His TinZiad AlazthaNo ratings yet
- Orthopaedic JournalDocument6 pagesOrthopaedic JournalZiad AlazthaNo ratings yet
- 008 BHJKVGHJXFHXDFHFDocument2 pages008 BHJKVGHJXFHXDFHFZiad AlazthaNo ratings yet
- 003 BuyuiguiohuioDocument2 pages003 BuyuiguiohuioZiad AlazthaNo ratings yet
- CekDocument1 pageCekZiad AlazthaNo ratings yet
- Trom BusDocument1 pageTrom BusZiad AlazthaNo ratings yet
- 003Document1 page003Ziad AlazthaNo ratings yet
- Cerebrospinal FluidDocument1 pageCerebrospinal FluidZiad AlazthaNo ratings yet
- Jurnal Reading Objectively Monitored Patching Regimens For Treatment of Amblyopia: Randomised TrialDocument1 pageJurnal Reading Objectively Monitored Patching Regimens For Treatment of Amblyopia: Randomised TrialZiad AlazthaNo ratings yet
- Jurnal Reading Objectively Monitored Patching Regimens For Treatment of Amblyopia: Randomised TrialDocument1 pageJurnal Reading Objectively Monitored Patching Regimens For Treatment of Amblyopia: Randomised TrialZiad AlazthaNo ratings yet
- Jurnal Reading Objectively Monitored Patching Regimens For Treatment of Amblyopia: Randomised TrialDocument1 pageJurnal Reading Objectively Monitored Patching Regimens For Treatment of Amblyopia: Randomised TrialZiad AlazthaNo ratings yet
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (120)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Make An Acknowledgment Receipt Showing A Full Payment of A Loan (Extinguishment by Payment)Document5 pagesMake An Acknowledgment Receipt Showing A Full Payment of A Loan (Extinguishment by Payment)Kim Cyrah Amor GerianNo ratings yet
- Espiloy COT DLP Math 5 Q2Document7 pagesEspiloy COT DLP Math 5 Q2Adeline Rose B. Espiloy100% (3)
- Heritage Halls HousingDocument3 pagesHeritage Halls HousingBrevan EllefsenNo ratings yet
- Kentucky National Guard MemorialDocument60 pagesKentucky National Guard MemorialCourier JournalNo ratings yet
- Management Final Paper 27062020 044941pm PDFDocument8 pagesManagement Final Paper 27062020 044941pm PDFHuzaifa KhanNo ratings yet
- Ch05 P24 Build A ModelDocument5 pagesCh05 P24 Build A ModelKatarína HúlekováNo ratings yet
- InTech-Molar Incisor Hypomineralization Morphological Aetiological Epidemiological and Clinical ConsiderationsDocument25 pagesInTech-Molar Incisor Hypomineralization Morphological Aetiological Epidemiological and Clinical ConsiderationsNeagu EmaNo ratings yet
- WIPO - ROMARIN - International Registration Details 20.3.2015Document4 pagesWIPO - ROMARIN - International Registration Details 20.3.2015Володимир ПриходченкоNo ratings yet
- A Room of One's Own EssayDocument3 pagesA Room of One's Own EssayPeter Hadley MahotiereNo ratings yet
- Nestle Philippines, Inc., v. PuedanDocument1 pageNestle Philippines, Inc., v. PuedanJoycee ArmilloNo ratings yet
- Beyond SyncretismDocument27 pagesBeyond Syncretismz miriamNo ratings yet
- Sample Thesis of Medical Record SystemDocument8 pagesSample Thesis of Medical Record Systemfj9dbfw4100% (1)
- CONCEPT DESIGN ALUM C.W SCHUCO ANDALUSIA Rev01.10.03.222Document118 pagesCONCEPT DESIGN ALUM C.W SCHUCO ANDALUSIA Rev01.10.03.222MoustafaNo ratings yet
- 1 Assignment-2Document8 pages1 Assignment-2abhiNo ratings yet
- The Global Workforce Crisis BCG PDFDocument28 pagesThe Global Workforce Crisis BCG PDFdakintaurNo ratings yet
- Stop TB Text Only 2012Document30 pagesStop TB Text Only 2012Ga B B OrlonganNo ratings yet
- Poultry: Professional Products For The Poultry IndustryDocument1 pagePoultry: Professional Products For The Poultry IndustryzoilaNo ratings yet
- Tugas Lk7 Dan 10 Garuda PancasilaDocument5 pagesTugas Lk7 Dan 10 Garuda PancasilaarifuddinNo ratings yet
- Chap3 Laterally Loaded Deep FoundationDocument46 pagesChap3 Laterally Loaded Deep Foundationtadesse habtieNo ratings yet
- The Achaeans (Also Called The "Argives" or "Danaans")Document3 pagesThe Achaeans (Also Called The "Argives" or "Danaans")Gian Paul JavierNo ratings yet
- WB TBA19 020vadvance PDFDocument266 pagesWB TBA19 020vadvance PDFTran Khang92% (12)
- Content-Area Instruction For Ells: Connecti Es ArchDocument15 pagesContent-Area Instruction For Ells: Connecti Es Archnickelt23No ratings yet
- Can Amitriptyline Makes You Feel More Awake Instead of DrowsyDocument3 pagesCan Amitriptyline Makes You Feel More Awake Instead of Drowsyteddypol100% (1)
- Carbon Trading-The Future Money Venture For IndiaDocument11 pagesCarbon Trading-The Future Money Venture For IndiaijsretNo ratings yet
- 2021 Community ReportDocument28 pages2021 Community Reportapi-309161587No ratings yet
- Deed of Donation BicycleDocument2 pagesDeed of Donation BicycleCharlemagne PapioNo ratings yet
- Finance Management Individual AssDocument33 pagesFinance Management Individual AssDevan MoroganNo ratings yet
- Unit 2 - Chapter 2Document20 pagesUnit 2 - Chapter 2Dennise Kate CabiedesNo ratings yet
- Grade 12 Mathematical Literacy: Question Paper 1 MARKS: 150 TIME: 3 HoursDocument53 pagesGrade 12 Mathematical Literacy: Question Paper 1 MARKS: 150 TIME: 3 HoursOfentse MothapoNo ratings yet
- Loan-Level Price Adjustment (LLPA) MatrixDocument8 pagesLoan-Level Price Adjustment (LLPA) MatrixLashon SpearsNo ratings yet