You might also like
- Protein Synthesis Race WorksheetDocument2 pagesProtein Synthesis Race WorksheetHugo FloresNo ratings yet
- Factors Affecting Enzyme ActionDocument18 pagesFactors Affecting Enzyme Actionanon_458882066No ratings yet
- Western Blotting PDFDocument17 pagesWestern Blotting PDFAna Dominique EspiaNo ratings yet
- Energy Balance On Separator (Correction)Document4 pagesEnergy Balance On Separator (Correction)nabilaNo ratings yet
- 04 Structure To FunctionDocument5 pages04 Structure To Function897291868100% (1)
- Human Chromosomes: An Illustrated Introduction to Human CytogeneticsFrom EverandHuman Chromosomes: An Illustrated Introduction to Human CytogeneticsRating: 5 out of 5 stars5/5 (1)
- Science 10 DLL Q3 W4Document6 pagesScience 10 DLL Q3 W4Lyno Rey100% (4)
- Amino Acid MCQ with AnswersDocument4 pagesAmino Acid MCQ with AnswersPrince AsanteNo ratings yet
- Protein FoldingDocument21 pagesProtein FoldingRONAK LASHKARINo ratings yet
- Biomolecules Chemistry AssignmentDocument28 pagesBiomolecules Chemistry AssignmentAnurag Singh Thakur67% (3)
- Ribosome Structure and FunctionDocument24 pagesRibosome Structure and FunctionMaliha JahanNo ratings yet
- Chapter 5Document10 pagesChapter 5Pee Wee Martini0% (1)
- Endoplasmic Reticulum and RibosomesDocument16 pagesEndoplasmic Reticulum and RibosomesRis habhNo ratings yet
- Tdnº3 103030Document3 pagesTdnº3 103030bachir zazaNo ratings yet
- 1583730232049-Endoplasmic ReticulumDocument8 pages1583730232049-Endoplasmic ReticulumGautamiNo ratings yet
- RIBOSOMESDocument6 pagesRIBOSOMESVidushi GuptaNo ratings yet
- The Nucleus and RibosomesDocument9 pagesThe Nucleus and RibosomestadiwaospencioNo ratings yet
- How The Ribosome Works American ScientistDocument6 pagesHow The Ribosome Works American ScientistDr Carlos Estrada VázquezNo ratings yet
- Cellular Compartments ExplainedDocument26 pagesCellular Compartments ExplainedAsim Bin ArshadNo ratings yet
- Ribosomes (Protein Factory) : Ribosome Rrna Rrna 40S 60SDocument4 pagesRibosomes (Protein Factory) : Ribosome Rrna Rrna 40S 60Ssajesh72No ratings yet
- Cell Ribosome: Tiny Protein FactoriesDocument5 pagesCell Ribosome: Tiny Protein FactoriesSyrusNo ratings yet
- RibosomeDocument14 pagesRibosomenadia riazNo ratings yet
- Ribosomes and Protein SynthesisDocument12 pagesRibosomes and Protein SynthesisSulfeekker AhamedNo ratings yet
- Ribosomes: Synonyms: Ribonucleoprotein (RNP) Factories, Palade Particles, Protein Factories, Claude'sDocument4 pagesRibosomes: Synonyms: Ribonucleoprotein (RNP) Factories, Palade Particles, Protein Factories, Claude'sNelecaupNo ratings yet
- Resume 293-305Document8 pagesResume 293-305Amirah NviNo ratings yet
- Ribosomes PPT NewDocument36 pagesRibosomes PPT NewVINOTHINI G100% (2)
- Ribosomal RnaDocument10 pagesRibosomal Rnaspdharanimaran001No ratings yet
- Ribosomal RNADocument18 pagesRibosomal RNAwilliam919No ratings yet
- 7 RibosomesDocument2 pages7 RibosomesSabina MaharjanNo ratings yet
- Ribosome Structure and Assembly: Durgeshwer Singh Department of Botany Mahatma Gandhi Central UniversityDocument12 pagesRibosome Structure and Assembly: Durgeshwer Singh Department of Botany Mahatma Gandhi Central Universityمحمّد نعیمNo ratings yet
- Ribozymes: Ribonuclease PDocument14 pagesRibozymes: Ribonuclease Pabash_u1No ratings yet
- Granular Endoplasmic ReticulumDocument2 pagesGranular Endoplasmic ReticulumSetumil PowerNo ratings yet
- RNA Structure and Functions in 40 CharactersDocument7 pagesRNA Structure and Functions in 40 CharacterssafemindNo ratings yet
- Ribosome and Protein Synthesis: By: Delfi Zurita NIM: RSA1C411009Document16 pagesRibosome and Protein Synthesis: By: Delfi Zurita NIM: RSA1C411009Atikatul MutmainahNo ratings yet
- Tema 3 - Ribosome - Gavrila Lucian-IonutDocument3 pagesTema 3 - Ribosome - Gavrila Lucian-IonutIonut ManolescuNo ratings yet
- The Ribosome Ribosomes A Re The Sites of Protein Synthesis in Both Prokaryotic and Eukaryotic CellsDocument2 pagesThe Ribosome Ribosomes A Re The Sites of Protein Synthesis in Both Prokaryotic and Eukaryotic CellsRatri MuktiNo ratings yet
- RIBOSOMESDocument10 pagesRIBOSOMESJorge BazánNo ratings yet
- LysosomesDocument29 pagesLysosomesBhavana ReddyNo ratings yet
- Cell NursingDocument83 pagesCell NursingRocKyRiazNo ratings yet
- Ribosome - Wikipedia 3Document2 pagesRibosome - Wikipedia 3Krishnamayi MRNo ratings yet
- Purification of Ribosomes From Human Cell Lines: Naskah ReviewDocument9 pagesPurification of Ribosomes From Human Cell Lines: Naskah ReviewFredho CasperNo ratings yet
- Distinguishing Characteristics: Key Takeaways: RibosomesDocument9 pagesDistinguishing Characteristics: Key Takeaways: RibosomesAnonymous yCpjZF1rFNo ratings yet
- Protein Synthesis (Translation) - Microbiology - OpenStaxDocument7 pagesProtein Synthesis (Translation) - Microbiology - OpenStaxAleksandra Sanja MartinovicNo ratings yet
- A-Endoplasmic Reticulum - Wrapping It Up: Rough and SmoothDocument9 pagesA-Endoplasmic Reticulum - Wrapping It Up: Rough and SmoothRahul PurwarNo ratings yet
- Functions of The Rough Endoplasmic Reticulum: Synthesis of Proteins On Membrane-Bound Versus Free RibosomesDocument7 pagesFunctions of The Rough Endoplasmic Reticulum: Synthesis of Proteins On Membrane-Bound Versus Free RibosomesDanie MorenoNo ratings yet
- Cell Organelles - Lec. 1Document27 pagesCell Organelles - Lec. 1Hamza KaisNo ratings yet
- Protein Translation Process ExplainedDocument20 pagesProtein Translation Process ExplainedFathiaNo ratings yet
- 3 CytoplasmDocument40 pages3 CytoplasmSHAZAIB ARSHIDNo ratings yet
- Ribosom 30s & 50sDocument3 pagesRibosom 30s & 50sFadhilah Putri Aulia BahriNo ratings yet
- Structure of RibosomesDocument15 pagesStructure of RibosomesSaba Parvin Haque100% (1)
- MolbioDocument4 pagesMolbioPrachi ShuklaNo ratings yet
- Ribozyme - WikipediaDocument61 pagesRibozyme - WikipediaBhavyaNo ratings yet
- Ribosomes Are Composed of Two Conserved SubunitsDocument4 pagesRibosomes Are Composed of Two Conserved Subunitsvaseem1981No ratings yet
- 2010 - Pulk - Ribosome Reactivation by Replacement of Damaged ProteinsDocument14 pages2010 - Pulk - Ribosome Reactivation by Replacement of Damaged ProteinsLai weng yu Lai weng yuNo ratings yet
- Biology NotesDocument42 pagesBiology NotesJoel ThompsonNo ratings yet
- Eukaryotic and Prokaryotic RibosomesDocument15 pagesEukaryotic and Prokaryotic RibosomesRININo ratings yet
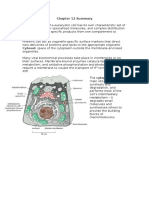
- Chapter 12 SummaryDocument7 pagesChapter 12 SummaryCharlotteNo ratings yet
- Biomolecules Chemistry AssignmentDocument27 pagesBiomolecules Chemistry AssignmentUvan VijayNo ratings yet
- Week 15 Lecture 560B On LineDocument9 pagesWeek 15 Lecture 560B On LineTheNourishedSproutNo ratings yet
- CellDocument8 pagesCellWhyL NificentNo ratings yet
- RibosomesDocument9 pagesRibosomesMeg Dianne V. CañedaNo ratings yet
- RibozymesDocument25 pagesRibozymesMohd JaheerNo ratings yet
- RibosomesDocument12 pagesRibosomesEnika baidenNo ratings yet
- Ribosom Kloroplas EMBJ 36 475Document12 pagesRibosom Kloroplas EMBJ 36 475NovyKedungWilutNo ratings yet
- Nucleic Acids Rna World Biotech ProjectDocument18 pagesNucleic Acids Rna World Biotech ProjectSakshi SinghNo ratings yet
- Plant Cell StructureDocument13 pagesPlant Cell Structureshamosa85.saNo ratings yet
- Archaeal Ribosomes: Structural and Functional AdaptationsDocument4 pagesArchaeal Ribosomes: Structural and Functional Adaptationsmanoj_rkl_070% (1)
- Er, Structure & Its FunctionsDocument20 pagesEr, Structure & Its FunctionsTalha AfzalNo ratings yet
- JFM Is The Participation of Local Community in The Managing ForestsDocument1 pageJFM Is The Participation of Local Community in The Managing ForestsKalyani Sawarkar SatoneNo ratings yet
- Mod3 PDFDocument65 pagesMod3 PDFRAKISHO WORLD100% (1)
- EiaDocument34 pagesEiaKalyani Sawarkar SatoneNo ratings yet
- Principles of Cloning, Vectors and Cloning StrategiesDocument93 pagesPrinciples of Cloning, Vectors and Cloning StrategiesEmmanuel OcanseyNo ratings yet
- What Are Transgenic PlantsDocument17 pagesWhat Are Transgenic PlantsKalyani Sawarkar SatoneNo ratings yet
- Mcqs Test: Biology Hssc-IDocument4 pagesMcqs Test: Biology Hssc-IZulfiqar ali Zulfiqar ali farhatNo ratings yet
- Biological Molecules: Types and Uses (39Document13 pagesBiological Molecules: Types and Uses (39fatema buhussainNo ratings yet
- Farmakologi FarmakodinamikDocument26 pagesFarmakologi FarmakodinamikMuhammad Nikko AomadaniNo ratings yet
- Enzyme Activity: BY Deeparubini P Snekha S MDocument20 pagesEnzyme Activity: BY Deeparubini P Snekha S MdeepaNo ratings yet
- Molecular Biology QuestionsDocument6 pagesMolecular Biology QuestionsVinay Kumar100% (1)
- MCQ AduaDocument4 pagesMCQ AduaAG KhanNo ratings yet
- Grade 12 Dna QuizDocument3 pagesGrade 12 Dna QuizDawnNo ratings yet
- Protein Sorting Nucleus & Mitochondrion PDFDocument8 pagesProtein Sorting Nucleus & Mitochondrion PDFHyunji KimNo ratings yet
- CH 3 Proteins Student PrintDocument23 pagesCH 3 Proteins Student PrintFazreen DzulkafliNo ratings yet
- Cloning and Expression - FLAG and 3xFLAG OverviewDocument3 pagesCloning and Expression - FLAG and 3xFLAG OverviewSigma-Aldrich100% (12)
- Overview of Post-Translational Modification AnalysisDocument2 pagesOverview of Post-Translational Modification AnalysisSteven4654No ratings yet
- Amino AcidsDocument6 pagesAmino AcidsJohannaNo ratings yet
- Especificaciones Nutritivas - Hyline RosDocument16 pagesEspecificaciones Nutritivas - Hyline RosPatriciaNo ratings yet
- Biosint de LipDocument24 pagesBiosint de LipdborghiNo ratings yet
- Nucleotides and Nucleic Acids: The Building Blocks of LifeDocument71 pagesNucleotides and Nucleic Acids: The Building Blocks of LifekalkidanNo ratings yet
- Protein Targeting: After This Chapter, You Should Be Able ToDocument12 pagesProtein Targeting: After This Chapter, You Should Be Able ToAmeena SherinNo ratings yet
- Hole's Human Anatomy and Physiology: Shier Butler LewisDocument12 pagesHole's Human Anatomy and Physiology: Shier Butler LewisTheophilus BaidooNo ratings yet
- Rescue of Folding Defects in ABC Transporters Using Pharmacological ChaperonesDocument7 pagesRescue of Folding Defects in ABC Transporters Using Pharmacological ChaperonesOwaisNo ratings yet
- MDSC1104 Tutorial on Hereditary Material and Gene ExpressionDocument2 pagesMDSC1104 Tutorial on Hereditary Material and Gene ExpressionJordan ConstantineNo ratings yet
- Pantoja, Ariel S. 1-Y2-12 Bs NursingDocument2 pagesPantoja, Ariel S. 1-Y2-12 Bs NursingChristopher GalivoNo ratings yet
- Learning Goal: Karen C. Timberlake © 2016 Pearson Education, IncDocument12 pagesLearning Goal: Karen C. Timberlake © 2016 Pearson Education, IncLleana TanNo ratings yet