You might also like
- OSCE Cheat SheetDocument3 pagesOSCE Cheat SheetazarsamaniNo ratings yet
- Cell Biology Test 3Document11 pagesCell Biology Test 3Carol HathawayNo ratings yet
- The Ten-Step Vaginal Hysterectomy - A Newer and Better ApproachDocument8 pagesThe Ten-Step Vaginal Hysterectomy - A Newer and Better ApproachqisthiaufaNo ratings yet
- Funda QuestionsDocument3 pagesFunda QuestionsKaye CorNo ratings yet
- A Runx-Cbf Β-Driven Enhancer Directs The Irf8 Dose-Dependent Lineage Choice Between Dcs And MonocytesDocument34 pagesA Runx-Cbf Β-Driven Enhancer Directs The Irf8 Dose-Dependent Lineage Choice Between Dcs And MonocytesD. S.No ratings yet
- HIV Entry and Its Inhibition by Bifunctional Antiviral ProteinsDocument18 pagesHIV Entry and Its Inhibition by Bifunctional Antiviral ProteinsmurielNo ratings yet
- CD Marker HandbookDocument47 pagesCD Marker HandbookSteph Vee100% (2)
- CD2BP2-GYF Interacts With The Spliceosomal Protein SMB - B' 1 (PDFDrive)Document31 pagesCD2BP2-GYF Interacts With The Spliceosomal Protein SMB - B' 1 (PDFDrive)Blink SNo ratings yet
- Cell Cycle & ApoptosisDocument42 pagesCell Cycle & ApoptosisRAHMA TRIYANAYNo ratings yet
- CD Markers HandbookDocument48 pagesCD Markers HandbookOmair Riaz100% (2)
- Cyclin Dependent Kinase: An IntroductionDocument35 pagesCyclin Dependent Kinase: An Introductionglreddy09No ratings yet
- The Cell Cycle: Sri DarmawatiDocument44 pagesThe Cell Cycle: Sri DarmawatiFahrum BurhanNo ratings yet
- CDC14 TreatiseDocument12 pagesCDC14 TreatiseasdfweafsplashNo ratings yet
- Stewart 2003Document7 pagesStewart 2003Arjun SNo ratings yet
- Cell Cycle SMDocument15 pagesCell Cycle SMOmphile DansonNo ratings yet
- Hereditary Cancer SyndromesDocument10 pagesHereditary Cancer SyndromesANA GABRIELA PEREZ VIDALNo ratings yet
- Loreto, M (2002) - Functional Cooperation Between C-CBL and Src-Like Adaptor Protein 2 in The Negative Regulation of T-Cell Receptor SignalingDocument15 pagesLoreto, M (2002) - Functional Cooperation Between C-CBL and Src-Like Adaptor Protein 2 in The Negative Regulation of T-Cell Receptor SignalingAnadahiNo ratings yet
- Cell Cycle Checkpoints As Therapeutic Targets: Zoe A. Stewart and Jennifer A. PietenpolDocument12 pagesCell Cycle Checkpoints As Therapeutic Targets: Zoe A. Stewart and Jennifer A. PietenpolDewi Sekar MiasihNo ratings yet
- Patterns of Cell Cycle Checkpoint Deregulation Associated With Intrinsic Molecular Subtypes of Human Breast Cancer CellsDocument12 pagesPatterns of Cell Cycle Checkpoint Deregulation Associated With Intrinsic Molecular Subtypes of Human Breast Cancer CellsErmiyas YeshitlaNo ratings yet
- Regulation of Hippo Pathway Through Mitogenic Growth Factors Via PI3K and PDK1Document18 pagesRegulation of Hippo Pathway Through Mitogenic Growth Factors Via PI3K and PDK1Verda Batool - 85817/TCHR/BMMCNo ratings yet
- Jurnal2 PDFDocument12 pagesJurnal2 PDFRINDA HERNISNo ratings yet
- 7.013 Exam 2 PDFDocument6 pages7.013 Exam 2 PDF15klaNo ratings yet
- 2017 Dendritic Cells in Cancer The Role RevisitedDocument9 pages2017 Dendritic Cells in Cancer The Role RevisitedAngela CandrevaNo ratings yet
- A Pilot Along With Power Soreness in The FacejpybgDocument2 pagesA Pilot Along With Power Soreness in The Facejpybgdealkevin60No ratings yet
- TMP 59 F3Document5 pagesTMP 59 F3FrontiersNo ratings yet
- Retroviral-Mediated Very-Long-Chain Fatty Acid Metabolism in Adrenoleukodystrophy FibroblastsDocument5 pagesRetroviral-Mediated Very-Long-Chain Fatty Acid Metabolism in Adrenoleukodystrophy FibroblastsmdquevedoNo ratings yet
- PDGF D and Renal Disease Yet Another One of Those.35Document2 pagesPDGF D and Renal Disease Yet Another One of Those.35Arista RachmaNo ratings yet
- Ji 2300122Document11 pagesJi 2300122api-716978431No ratings yet
- PIIS0021925820312412Document10 pagesPIIS0021925820312412Jose FernandezNo ratings yet
- BCH 323 Tutorial 4 2019 Memo 14th October 2019Document4 pagesBCH 323 Tutorial 4 2019 Memo 14th October 2019Nosibusiso KhaliphaNo ratings yet
- Decker 1Document9 pagesDecker 1Katarina Kaca GacevicNo ratings yet
- Cell Cycle: RNA, Protein Mitosis, CytokinesisDocument4 pagesCell Cycle: RNA, Protein Mitosis, Cytokinesisblueradish450No ratings yet
- MCQ in BioDocument7 pagesMCQ in BioGiovanni Roccaforte100% (1)
- tmpCF25 TMPDocument10 pagestmpCF25 TMPFrontiersNo ratings yet
- The Cell Cycle in Stem Cell Proliferation, Pluripotency and DifferentiationDocument8 pagesThe Cell Cycle in Stem Cell Proliferation, Pluripotency and DifferentiationoksanitaposadasposaditasNo ratings yet
- J. Biol. Chem.-2000-Del Peso-27205-11Document7 pagesJ. Biol. Chem.-2000-Del Peso-27205-11Thomas IngallsNo ratings yet
- Ficha de ExercíciosDocument10 pagesFicha de ExercíciosEltinNo ratings yet
- Analysis of CD16þCD56dim NK Cells From CLL Patients Evidence Supporting ADocument9 pagesAnalysis of CD16þCD56dim NK Cells From CLL Patients Evidence Supporting Aimmunologie cellulaireNo ratings yet
- KA P:S - The Lazy OCD VersionDocument9 pagesKA P:S - The Lazy OCD VersionsaialamNo ratings yet
- Vockerodt Et Al-2008-The Journal of PathologyDocument10 pagesVockerodt Et Al-2008-The Journal of PathologyBeatrice FacchiniNo ratings yet
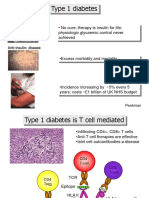
- Type 1 Diabetes: - No Cure Therapy Is Insulin For Life Physiologic Glycaemic Control Never AchievedDocument39 pagesType 1 Diabetes: - No Cure Therapy Is Insulin For Life Physiologic Glycaemic Control Never AchievedHimawan SartonoNo ratings yet
- Complement SystemDocument38 pagesComplement Systemprateek gangwaniNo ratings yet
- Art:10.1186/1747 1028 4 2 - APC - cdh1Document7 pagesArt:10.1186/1747 1028 4 2 - APC - cdh1TarunSreerajNo ratings yet
- Lab ProgressDocument37 pagesLab Progressirine14omegaNo ratings yet
- 2018, Nature, CHR2Document34 pages2018, Nature, CHR2OLGA MARÍNNo ratings yet
- 2018 Lecture10Document35 pages2018 Lecture10Adam Bryant PoonawalaNo ratings yet
- Yen & Jaffe's Reproductive Endocrinology (PDFDrive) - Pages-8Document17 pagesYen & Jaffe's Reproductive Endocrinology (PDFDrive) - Pages-8viramaharsaNo ratings yet
- 2002; Two DBLγ Subtypes Are Commonly Expressed by Placental Isolates of Plasmodium FalciparumDocument10 pages2002; Two DBLγ Subtypes Are Commonly Expressed by Placental Isolates of Plasmodium FalciparumSethawud ChaikitgosiyakulNo ratings yet
- 15-Deoxy - Prostaglandin J Induces IL-8 and GM-CSF in A Human Airway Epithelial Cell Line (NCI-H)Document7 pages15-Deoxy - Prostaglandin J Induces IL-8 and GM-CSF in A Human Airway Epithelial Cell Line (NCI-H)Roy GoldenNo ratings yet
- Mammalian Rho GTPases New Insights Into Their Functions From in Vivo Studies 05Document12 pagesMammalian Rho GTPases New Insights Into Their Functions From in Vivo Studies 05Sonia Barbosa CornelioNo ratings yet
- T Cell Activation and Signaling: TCR TalinDocument8 pagesT Cell Activation and Signaling: TCR TalinMst Monira KhatonNo ratings yet
- F22 MCB 2050 Lecture 4 - Gene RegulationDocument29 pagesF22 MCB 2050 Lecture 4 - Gene RegulationNO VIDEOSNo ratings yet
- The Polycomb Group Protein Yaf2 Regulates The Pluripotency of Embryonic Stem Cells in A Phosphorylation-Dependent MannerDocument13 pagesThe Polycomb Group Protein Yaf2 Regulates The Pluripotency of Embryonic Stem Cells in A Phosphorylation-Dependent MannerSHUMETNo ratings yet
- Lodish8e Ch15 TestBankDocument17 pagesLodish8e Ch15 TestBanksyg0302No ratings yet
- IFN e VEGF-CDocument11 pagesIFN e VEGF-CThádia AraújoNo ratings yet
- Ochiai Et Al 2024 Accelerated Plasma Cell Differentiation in Bach2 Deficient Mouse B Cells Is Caused by Altered Irf4Document18 pagesOchiai Et Al 2024 Accelerated Plasma Cell Differentiation in Bach2 Deficient Mouse B Cells Is Caused by Altered Irf4Daniel DavidNo ratings yet
- Calderilla-Barbosa Et Al-2006-Journal of Neurochemistry PDFDocument10 pagesCalderilla-Barbosa Et Al-2006-Journal of Neurochemistry PDFLuis CalderillaNo ratings yet
- 7.02 Recombinant DNA Methods Spring 2005 Exam Study Questions Answer KeyDocument10 pages7.02 Recombinant DNA Methods Spring 2005 Exam Study Questions Answer KeyFCiênciasNo ratings yet
- Duyk 1990Document5 pagesDuyk 1990Vishnu Sai MokshagnaNo ratings yet
- Expression Profile of Intrinsic SubtypesDocument63 pagesExpression Profile of Intrinsic SubtypesBikash Ranjan SamalNo ratings yet
- Neutro PhilsDocument39 pagesNeutro PhilsAjamalSingh BhayalNo ratings yet
- Apoptosis: Dr. Bimal PatelDocument22 pagesApoptosis: Dr. Bimal PatelBimal PatelNo ratings yet
- Itchs 2012Document2,486 pagesItchs 2012Vishal BhardwajNo ratings yet
- Cranial NervesDocument67 pagesCranial NervesBahaa ShaabanNo ratings yet
- Endocrine Pharmacology PDFDocument45 pagesEndocrine Pharmacology PDFNur TzNo ratings yet
- Unit 1-Heat CVD Lungs CF QuestionsDocument73 pagesUnit 1-Heat CVD Lungs CF Questionsareyouthere9267% (3)
- EPIDIMIOLOGYDocument13 pagesEPIDIMIOLOGYPeter MainyaNo ratings yet
- Philippine National Standard: Code of Slaughtering Practices For GoatsDocument13 pagesPhilippine National Standard: Code of Slaughtering Practices For GoatsJRSA SAALNo ratings yet
- HematotoraksDocument15 pagesHematotorakssafanj12No ratings yet
- Myringotomy PET PT INFODocument3 pagesMyringotomy PET PT INFOdhiahabsyiNo ratings yet
- Effect of TetanospasminDocument23 pagesEffect of Tetanospasminaparna ranjith markoseNo ratings yet
- BS (4 Ye Ears) For A Affiliated Co Olleges: Course CDocument2 pagesBS (4 Ye Ears) For A Affiliated Co Olleges: Course CBilal AhmadNo ratings yet
- Yin Yang PrimerDocument66 pagesYin Yang PrimerAnais100% (2)
- Canine Environmental EnrichmentDocument3 pagesCanine Environmental EnrichmentThamil ArasanNo ratings yet
- College of Veterinary and Animal Science NavaniaDocument16 pagesCollege of Veterinary and Animal Science NavaniaNeerajNo ratings yet
- Vocabulary Unit 4 PrintableDocument2 pagesVocabulary Unit 4 PrintableLuis EnriqueNo ratings yet
- 5th Year Pediatric Exam 2016Document14 pages5th Year Pediatric Exam 2016pal_pal_palNo ratings yet
- Skeletal System PowerpointDocument40 pagesSkeletal System PowerpointLee KaiYangNo ratings yet
- MiliariaDocument6 pagesMiliariaZyzah HasibuanNo ratings yet
- How To Increase Breast SizeDocument7 pagesHow To Increase Breast SizeAndrewJonathanNo ratings yet
- Introduction To ParasitologyDocument79 pagesIntroduction To ParasitologyLeeShauran100% (7)
- 3776011564Document120 pages3776011564chaido1100% (4)
- Hypertensive Thalamic Hemorrhage: Analysis of Short-Term OutcomeDocument6 pagesHypertensive Thalamic Hemorrhage: Analysis of Short-Term OutcomeErwin Chiquete, MD, PhDNo ratings yet
- Lesson 9 Questions & AnswersDocument2 pagesLesson 9 Questions & AnswersAnonymous nulAkhlY100% (2)
- 12 Week USMLE Schedule PDFDocument5 pages12 Week USMLE Schedule PDFChristianNo ratings yet
- Introduction About SelfDocument27 pagesIntroduction About SelfsimonjosanNo ratings yet
- HM 12 38078v1 WW Ruby Brochure 8.5x11 100112Document8 pagesHM 12 38078v1 WW Ruby Brochure 8.5x11 100112vijayramaswamyNo ratings yet
- Buser Et Al-2017-Periodontology 2000Document19 pagesBuser Et Al-2017-Periodontology 2000andreita_loppNo ratings yet
- Date 03/aug/2023 07:32AM Unit Bio Ref Interval: Laboratory Investigation ReportDocument12 pagesDate 03/aug/2023 07:32AM Unit Bio Ref Interval: Laboratory Investigation ReportRishabh GuptaNo ratings yet