You might also like
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (121)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- EX200Document7 pagesEX200shubbyNo ratings yet
- A Teachers' Journey: Phenomenological Study On The Puritive Behavioral Standards of Students With Broken FamilyDocument11 pagesA Teachers' Journey: Phenomenological Study On The Puritive Behavioral Standards of Students With Broken FamilyNova Ariston100% (2)
- Iec Codes PDFDocument257 pagesIec Codes PDFAkhil AnumandlaNo ratings yet
- Priest, Graham - The Logic of The Catuskoti (2010)Document31 pagesPriest, Graham - The Logic of The Catuskoti (2010)Alan Ruiz100% (1)
- Case Report: A Novel, Fixed Chairside Space MaintainerDocument4 pagesCase Report: A Novel, Fixed Chairside Space MaintainerEcha VavataNo ratings yet
- The Effectiveness of Ethanolic Extract of Moringa Leaves (Moringa Oleifera Lam.) Gel On The Wound Healing Process of The Rat's PalateDocument6 pagesThe Effectiveness of Ethanolic Extract of Moringa Leaves (Moringa Oleifera Lam.) Gel On The Wound Healing Process of The Rat's PalateEcha VavataNo ratings yet
- Antibacterial Effect and Healing Potential of TopiDocument4 pagesAntibacterial Effect and Healing Potential of TopiEcha VavataNo ratings yet
- 13.D18 805 Inne Suhernasasmita2Document12 pages13.D18 805 Inne Suhernasasmita2Echa VavataNo ratings yet
- Khus Bu and Satyam Dental Caries ReviewDocument8 pagesKhus Bu and Satyam Dental Caries Reviewmeena syedNo ratings yet
- The Effectiveness of Ethanolic Extract of Moringa Leaves (Moringa Oleifera Lam.) Gel On The Wound Healing Process of The Rat's PalateDocument6 pagesThe Effectiveness of Ethanolic Extract of Moringa Leaves (Moringa Oleifera Lam.) Gel On The Wound Healing Process of The Rat's PalateEcha VavataNo ratings yet
- Abouneel 2014Document14 pagesAbouneel 2014Echa VavataNo ratings yet
- Abouneel 2014Document14 pagesAbouneel 2014Echa VavataNo ratings yet
- Health & Place: Jane Yeonjae Lee, Robin A. Kearns, Wardlow FriesenDocument8 pagesHealth & Place: Jane Yeonjae Lee, Robin A. Kearns, Wardlow FriesenEcha VavataNo ratings yet
- Gender Differences in The Association Between Tooth Loss and Obesity Among Older Adults in BrazilDocument9 pagesGender Differences in The Association Between Tooth Loss and Obesity Among Older Adults in BrazilEcha VavataNo ratings yet
- Experimental and Toxicologic PathologyDocument5 pagesExperimental and Toxicologic PathologyEcha VavataNo ratings yet
- Barrowman Et Al-2010-Australian Dental JournalDocument5 pagesBarrowman Et Al-2010-Australian Dental JournalEcha VavataNo ratings yet
- Hypoxia Enhances Periodontal Ligament Stem Cell Proliferation Via The Mapk Signaling Pathway PDFDocument10 pagesHypoxia Enhances Periodontal Ligament Stem Cell Proliferation Via The Mapk Signaling Pathway PDFEcha VavataNo ratings yet
- Time Table Clinical Dental Science Ii 2018/2019: Hari/ Tanggal JAM KegiatanDocument14 pagesTime Table Clinical Dental Science Ii 2018/2019: Hari/ Tanggal JAM KegiatanEcha VavataNo ratings yet
- CalendulaDocument13 pagesCalendulamery 21No ratings yet
- Barrowman Et Al-2010-Australian Dental JournalDocument5 pagesBarrowman Et Al-2010-Australian Dental JournalEcha VavataNo ratings yet
- G WH Histo Rinastiti2006 PDFDocument5 pagesG WH Histo Rinastiti2006 PDFEcha VavataNo ratings yet
- Crabs HellDocument6 pagesCrabs HellEcha VavataNo ratings yet
- CalendulaDocument13 pagesCalendulamery 21No ratings yet
- UnDocument3 pagesUnEcha VavataNo ratings yet
- Crabs HellDocument6 pagesCrabs HellEcha VavataNo ratings yet
- We Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsDocument23 pagesWe Are Intechopen, The World'S Leading Publisher of Open Access Books Built by Scientists, For ScientistsEcha VavataNo ratings yet
- CalendulaDocument13 pagesCalendulamery 21No ratings yet
- Microbiological Research 20120172129Document10 pagesMicrobiological Research 20120172129Echa VavataNo ratings yet
- Benjamin Franklin - The Indian Treaties (1938)Document450 pagesBenjamin Franklin - The Indian Treaties (1938)Spiritu SanctoNo ratings yet
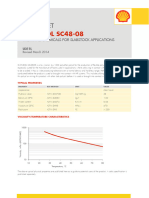
- Caradol sc48 08Document2 pagesCaradol sc48 08GİZEM DEMİRNo ratings yet
- William Hallett - BiographyDocument2 pagesWilliam Hallett - Biographyapi-215611511No ratings yet
- Second Conditional Conversation QuestionsDocument2 pagesSecond Conditional Conversation QuestionsEdith Salomé PinosNo ratings yet
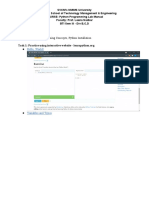
- Week - 2 Lab - 1 - Part I Lab Aim: Basic Programming Concepts, Python InstallationDocument13 pagesWeek - 2 Lab - 1 - Part I Lab Aim: Basic Programming Concepts, Python InstallationSahil Shah100% (1)
- Lesson 1: Composition: Parts of An EggDocument22 pagesLesson 1: Composition: Parts of An Eggjohn michael pagalaNo ratings yet
- 1 PBDocument7 pages1 PBIndah Purnama TaraNo ratings yet
- 300u Specs Diodo 300 Amps. 25 Dolares RadiosurtidoraDocument6 pages300u Specs Diodo 300 Amps. 25 Dolares RadiosurtidorarepelindNo ratings yet
- Java Complete Collection FrameworkDocument28 pagesJava Complete Collection FrameworkkhushivanshNo ratings yet
- BÀI TẬP LESSON 7. CÂU BỊ ĐỘNG 1Document4 pagesBÀI TẬP LESSON 7. CÂU BỊ ĐỘNG 1Yến Vy TrầnNo ratings yet
- Microfinance Ass 1Document15 pagesMicrofinance Ass 1Willard MusengeyiNo ratings yet
- The Comma Rules Conversion 15 SlidesDocument15 pagesThe Comma Rules Conversion 15 SlidesToh Choon HongNo ratings yet
- Ti05001 PDFDocument1 pageTi05001 PDFanggieNo ratings yet
- Table of Specification 1st QDocument5 pagesTable of Specification 1st QVIRGILIO JR FABINo ratings yet
- Lancru hzj105 DieselDocument2 pagesLancru hzj105 DieselMuhammad MasdukiNo ratings yet
- Final Test Level 7 New Format 2019Document3 pagesFinal Test Level 7 New Format 2019fabian serranoNo ratings yet
- Daewoo 710B PDFDocument59 pagesDaewoo 710B PDFbgmentNo ratings yet
- June 2017 (IAL) MS - Unit 1 Edexcel Physics A-LevelDocument16 pagesJune 2017 (IAL) MS - Unit 1 Edexcel Physics A-LevelNyraStardollNo ratings yet
- RN42Document26 pagesRN42tenminute1000No ratings yet
- Algorithms For Automatic Modulation Recognition of Communication Signals-Asoke K, Nandi, E.E AzzouzDocument6 pagesAlgorithms For Automatic Modulation Recognition of Communication Signals-Asoke K, Nandi, E.E AzzouzGONGNo ratings yet
- X - WORMWOOD EVENT IMMEDIATE - Paranormal - 4chanDocument7 pagesX - WORMWOOD EVENT IMMEDIATE - Paranormal - 4chanAnonymous dIjB7XD8ZNo ratings yet
- Tyler Nugent ResumeDocument3 pagesTyler Nugent Resumeapi-315563616No ratings yet
- Community Architecture Concept PDFDocument11 pagesCommunity Architecture Concept PDFdeanNo ratings yet
- Academic Socialization and Its Effects On Academic SuccessDocument2 pagesAcademic Socialization and Its Effects On Academic SuccessJustin LargoNo ratings yet
- Flow of FoodDocument2 pagesFlow of FoodGenevaNo ratings yet
- Sainik School Balachadi: Name-Class - Roll No - Subject - House - Assigned byDocument10 pagesSainik School Balachadi: Name-Class - Roll No - Subject - House - Assigned byPagalNo ratings yet