You might also like
- Reverse Underlying Cause of Degenerative Aging - Life ExtensionDocument15 pagesReverse Underlying Cause of Degenerative Aging - Life ExtensionJoshNo ratings yet
- Fast Facts pour les patients: Les troubles d'oxydation des acides gras à chaîne longueFrom EverandFast Facts pour les patients: Les troubles d'oxydation des acides gras à chaîne longueNo ratings yet
- AMPK - Metabolism & Insulin + Ways To Activate - SelfHackedDocument17 pagesAMPK - Metabolism & Insulin + Ways To Activate - SelfHackedJoshNo ratings yet
- The 5 Stages of Intermittent Fasting - LIFE Apps - LIVE and LEARNDocument8 pagesThe 5 Stages of Intermittent Fasting - LIFE Apps - LIVE and LEARNPetra CtiborNo ratings yet
- NIH Public Access: Author ManuscriptDocument34 pagesNIH Public Access: Author Manuscriptcamilamadrid01No ratings yet
- AMP-activated Protein Kinase Is Activated As A Consequence J. Biol. Chem.-2008-Gauthier-16514-24Document12 pagesAMP-activated Protein Kinase Is Activated As A Consequence J. Biol. Chem.-2008-Gauthier-16514-24evanconstantine77No ratings yet
- Glycogen QuizDocument2 pagesGlycogen QuizMohammed Elsaadi100% (1)
- HW01. Drug Reverses ObesityDocument1 pageHW01. Drug Reverses ObesityFlaviaNo ratings yet
- Regulat and ATP - TriTIme MagazineDocument3 pagesRegulat and ATP - TriTIme MagazineBiologicalMed99100% (1)
- Mitochondrial Medicine - ATP and RegulatDocument3 pagesMitochondrial Medicine - ATP and RegulatBiologicalMed99No ratings yet
- BF - v3.62 DRAFT RC1.5 ProtocolDocument48 pagesBF - v3.62 DRAFT RC1.5 Protocolmbw12294No ratings yet
- Lacture 5Document116 pagesLacture 5Ahmed Makky JafarNo ratings yet
- Journal Club: Paolo Dametto January 2013Document42 pagesJournal Club: Paolo Dametto January 2013Олег ЕвсюковNo ratings yet
- Running Head: Enzymology & Metabolism 1Document20 pagesRunning Head: Enzymology & Metabolism 1Jen ReyNo ratings yet
- 5 Stages of FastingDocument7 pages5 Stages of FastingMahdiNo ratings yet
- E. Estudiante - Reading Flow-Chart CompletionDocument2 pagesE. Estudiante - Reading Flow-Chart CompletionAna Maria CuartasNo ratings yet
- Metabolic Homeostasis: Regulation of Appetite, Energy Expenditure, Body Weight Unit-IVDocument26 pagesMetabolic Homeostasis: Regulation of Appetite, Energy Expenditure, Body Weight Unit-IVSherin SunnyNo ratings yet
- Academic Reading Sample Task Flowchart Completion Selecting Words From The Text TaskDocument2 pagesAcademic Reading Sample Task Flowchart Completion Selecting Words From The Text Taskharman randhawaNo ratings yet
- MAXGXL ProductDocument24 pagesMAXGXL ProductaprildelacuraNo ratings yet
- Recognizingandmanaginga Metaboliccrisis: Peter R. Baker IiDocument15 pagesRecognizingandmanaginga Metaboliccrisis: Peter R. Baker IiSara RamírezNo ratings yet
- Biological Theories of AgingDocument29 pagesBiological Theories of AgingdulceRN67% (3)
- Curcumin Effect On ObesityDocument7 pagesCurcumin Effect On ObesityAldo MataNo ratings yet
- Reversing Immunodeficiency PDFDocument8 pagesReversing Immunodeficiency PDFsorinNo ratings yet
- Ampk and The Adaptation To ExerciseDocument3 pagesAmpk and The Adaptation To ExerciseAndrés ChávezNo ratings yet
- Gardner ('18) - A Big Blog Post On Acetyl-L-Carnitine Benefits and Side EffectsDocument4 pagesGardner ('18) - A Big Blog Post On Acetyl-L-Carnitine Benefits and Side Effectsaida-69No ratings yet
- Endocrine System - MCAT Biology 5Document9 pagesEndocrine System - MCAT Biology 5Saran p sNo ratings yet
- ALCARDocument32 pagesALCARhydrogonianNo ratings yet
- Fasting and Low Carb InfoDocument4 pagesFasting and Low Carb InfoKatrin NovaNo ratings yet
- System For Heavy Lifting and Strenuous ActivitiesDocument7 pagesSystem For Heavy Lifting and Strenuous ActivitiesMaryAnnAnabeNo ratings yet
- Notes On EnzymologyDocument23 pagesNotes On EnzymologyCalvin Yeow-kuan ChongNo ratings yet
- Refined Sugar PDFDocument10 pagesRefined Sugar PDFFalk WendtNo ratings yet
- Effects of Intermittent Fasting On Health, Aging, and DiseaseDocument11 pagesEffects of Intermittent Fasting On Health, Aging, and DiseaseGonzalo TabaresNo ratings yet
- Lung Cancer and ALK F1174LDocument4 pagesLung Cancer and ALK F1174LAnandNo ratings yet
- Nutrigenetics & Fatty Acid Metabolism: Genes Analysed in ThisDocument21 pagesNutrigenetics & Fatty Acid Metabolism: Genes Analysed in ThisPetra JobovaNo ratings yet
- Insulin's Mechanism of ActionDocument14 pagesInsulin's Mechanism of ActionKishore ChinnaNo ratings yet
- Colostrum-LD AntiAging BenefitsDocument6 pagesColostrum-LD AntiAging BenefitsAntonio CarneiroNo ratings yet
- Immunocal Platinum Road To Good HealthDocument1 pageImmunocal Platinum Road To Good Healthapi-320558321No ratings yet
- Anti Aging Benefits of Bovine Colostrum Doug WyattDocument6 pagesAnti Aging Benefits of Bovine Colostrum Doug WyattFuzail NaseerNo ratings yet
- Autophagy & TomorigenisisDocument22 pagesAutophagy & TomorigenisisKirk CobainNo ratings yet
- Benfotiamine FALLDocument14 pagesBenfotiamine FALLzimkoNo ratings yet
- Plant Based Ketogenic Diet For CancerDocument25 pagesPlant Based Ketogenic Diet For CancerMNo ratings yet
- Glycine Reverses Aging in CellsDocument9 pagesGlycine Reverses Aging in CellsCarlos RodriguezNo ratings yet
- AMPK Modulates Tissue and Organismal Aging in A Non-Cell-Autonomous MannerDocument15 pagesAMPK Modulates Tissue and Organismal Aging in A Non-Cell-Autonomous MannerPilar Bravo SalasNo ratings yet
- Nutritional Support in Intensive Care Unit (ICU) PatientsDocument20 pagesNutritional Support in Intensive Care Unit (ICU) PatientssarahNo ratings yet
- The Scientific Basis For Metabolic Typing® - Part 2: The Autonomic Nervous System (ANS)Document3 pagesThe Scientific Basis For Metabolic Typing® - Part 2: The Autonomic Nervous System (ANS)jayjonbeachNo ratings yet
- Mechanism of Activation of The Cdk2Document3 pagesMechanism of Activation of The Cdk2MUHAMMAD ARIFFUDINNo ratings yet
- Recent Findings On NN-Dimethylgycine (DMG)Document9 pagesRecent Findings On NN-Dimethylgycine (DMG)K AnjaliNo ratings yet
- tp1 - EndoDocument5 pagestp1 - EndoRahadiyan HadinataNo ratings yet
- Chris Masterjohn Book DemoDocument15 pagesChris Masterjohn Book Demojijibiji24100% (1)
- Cosmeceutical PeptidesDocument7 pagesCosmeceutical PeptidesharborNo ratings yet
- NAM Tutorial Glycogen Storage DiseaseDocument2 pagesNAM Tutorial Glycogen Storage DiseaseAlzr FsNo ratings yet
- Adipokinas AntiinflamatoriasDocument8 pagesAdipokinas AntiinflamatoriasArturo PerezNo ratings yet
- Summary of Fat or FuelDocument6 pagesSummary of Fat or FuellengwaNo ratings yet
- Nutrients 11 02923 v2Document18 pagesNutrients 11 02923 v2Manaloe90No ratings yet
- UAS KARBOHIDRAT 2-DikonversiDocument18 pagesUAS KARBOHIDRAT 2-DikonversiumayaNo ratings yet
- Enzymes Article - CORRECTED VersionDocument2 pagesEnzymes Article - CORRECTED Versionaky13rxNo ratings yet
- Regulation of MetabolismDocument56 pagesRegulation of MetabolismdewkmcNo ratings yet
- AMPc y MetaboismoDocument8 pagesAMPc y MetaboismoEmilio MoncadaNo ratings yet
- ExamDocument36 pagesExamsnarkynurse with stetoscope0% (1)
- Diagnostic, Color of Smoke From Tailpipe - Smoking-Vehicle-NotificationDocument2 pagesDiagnostic, Color of Smoke From Tailpipe - Smoking-Vehicle-NotificationgstrohlNo ratings yet
- More Clues, and Questions, in 1961 Crash That Killed Dag HammarskjoldDocument3 pagesMore Clues, and Questions, in 1961 Crash That Killed Dag HammarskjoldgstrohlNo ratings yet
- List of Active Substances Approved For Use in Organic FarmingDocument1 pageList of Active Substances Approved For Use in Organic FarminggstrohlNo ratings yet
- White Liberals Dumb Themselves Down When They Speak To Black People, A New Study ContendsDocument14 pagesWhite Liberals Dumb Themselves Down When They Speak To Black People, A New Study ContendsgstrohlNo ratings yet
- White Liberals Dumb Themselves Down When They Speak To Black People, A New Study ContendsDocument14 pagesWhite Liberals Dumb Themselves Down When They Speak To Black People, A New Study ContendsgstrohlNo ratings yet
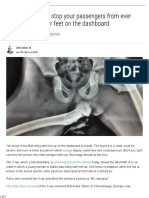
- X-Ray Vividly Shows Why You Shouldn't Ride With Feet On Car DashboardDocument3 pagesX-Ray Vividly Shows Why You Shouldn't Ride With Feet On Car DashboardgstrohlNo ratings yet
- More Clues, and Questions, in 1961 Crash That Killed Dag HammarskjoldDocument3 pagesMore Clues, and Questions, in 1961 Crash That Killed Dag HammarskjoldgstrohlNo ratings yet
- How To Buy Used Cars Cheap (Good Information To Know) !Document12 pagesHow To Buy Used Cars Cheap (Good Information To Know) !gstrohlNo ratings yet
- Decision Matrix Analysis - Decision-Making Skills FromDocument3 pagesDecision Matrix Analysis - Decision-Making Skills FromgstrohlNo ratings yet
- DecisionMatrixAnalysisWorksheet PDFDocument1 pageDecisionMatrixAnalysisWorksheet PDFLeo HutriNo ratings yet
- Tricks To Protect Your Eyes in Front of The PCDocument12 pagesTricks To Protect Your Eyes in Front of The PCKeyur P FNo ratings yet
- White Liberals Dumb Themselves Down When They Speak To Black People, A New Study ContendsDocument14 pagesWhite Liberals Dumb Themselves Down When They Speak To Black People, A New Study ContendsgstrohlNo ratings yet
- New Method To Slow Brain Aging by William Faloon and Cynthia RichardsDocument13 pagesNew Method To Slow Brain Aging by William Faloon and Cynthia RichardsgstrohlNo ratings yet
- Brain-Restoring Impact of Magnesium L-Threonate by Susan GoldscheinDocument7 pagesBrain-Restoring Impact of Magnesium L-Threonate by Susan GoldscheingstrohlNo ratings yet
- Can We Control Aging, Disease and Death by Saul KentDocument5 pagesCan We Control Aging, Disease and Death by Saul KentgstrohlNo ratings yet
- Cobra Breath TechniqueDocument2 pagesCobra Breath TechniqueJames Driscoll94% (16)
- Outwitting Our Aging Brain by William FaloonDocument18 pagesOutwitting Our Aging Brain by William FaloongstrohlNo ratings yet
- Brain-Restoring Impact of Magnesium L-Threonate by Susan GoldscheinDocument7 pagesBrain-Restoring Impact of Magnesium L-Threonate by Susan GoldscheingstrohlNo ratings yet
- Natural Compounds Remove Aging Cells by Sherrie OppenheimerDocument9 pagesNatural Compounds Remove Aging Cells by Sherrie OppenheimergstrohlNo ratings yet
- New Method To Slow Brain Aging by William Faloon and Cynthia RichardsDocument13 pagesNew Method To Slow Brain Aging by William Faloon and Cynthia RichardsgstrohlNo ratings yet
- Testosterone Replacement Therapy, A Recipe For Success - John Crisler, D.O.Document8 pagesTestosterone Replacement Therapy, A Recipe For Success - John Crisler, D.O.gstrohlNo ratings yet
- Cosmic Cobra Breath PDFDocument10 pagesCosmic Cobra Breath PDFgstrohlNo ratings yet
- Nikken MX2R Device, Pulsed Magnetic Field Therapy in Refractory CarpalDocument6 pagesNikken MX2R Device, Pulsed Magnetic Field Therapy in Refractory CarpalgstrohlNo ratings yet
- Open Angle Glaucoma - Epidemiology, Pathogenesis and PreventionDocument12 pagesOpen Angle Glaucoma - Epidemiology, Pathogenesis and PreventiongstrohlNo ratings yet
- Future of Product DesignDocument31 pagesFuture of Product DesignpericoNo ratings yet
- Bio Identical Hormone Replacement Therapy e BookDocument4 pagesBio Identical Hormone Replacement Therapy e BookgstrohlNo ratings yet
- Payments Industry Explained - The Trends Creating New Winners and Losers in The Card-Processing EcosystemDocument2 pagesPayments Industry Explained - The Trends Creating New Winners and Losers in The Card-Processing EcosystemgstrohlNo ratings yet
- eBook-How To Build A BrandDocument40 pageseBook-How To Build A Brandshlokshah2006No ratings yet
- eBook-How To Build A BrandDocument40 pageseBook-How To Build A Brandshlokshah2006No ratings yet
- Facultybooklet2011-2012final - 006 (Unlocked by WWW - Freemypdf.com)Document199 pagesFacultybooklet2011-2012final - 006 (Unlocked by WWW - Freemypdf.com)kalam19892209No ratings yet
- Travelstart Ticket (ZA10477979) PDFDocument2 pagesTravelstart Ticket (ZA10477979) PDFMatthew PretoriusNo ratings yet
- CAMEL Model With Detailed Explanations and Proper FormulasDocument4 pagesCAMEL Model With Detailed Explanations and Proper FormulasHarsh AgarwalNo ratings yet
- Yojananov 2021Document67 pagesYojananov 2021JackNo ratings yet
- DescriptiveDocument1 pageDescriptiveRizqa Anisa FadhilahNo ratings yet
- Starbucks Progressive Web App: Case StudyDocument2 pagesStarbucks Progressive Web App: Case StudyYesid SuárezNo ratings yet
- TOR AND SCOPING Presentation SlidesDocument23 pagesTOR AND SCOPING Presentation SlidesSRESTAA BHATTARAINo ratings yet
- Introduction To History AnswerDocument3 pagesIntroduction To History AnswerLawrence De La RosaNo ratings yet
- Johari WindowDocument7 pagesJohari WindowSarthak Priyank VermaNo ratings yet
- Data Structures and Algorithms SyllabusDocument9 pagesData Structures and Algorithms SyllabusBongbong GalloNo ratings yet
- PDFDocument3 pagesPDFvaliNo ratings yet
- Old Highland Park Baptist Church E01Document74 pagesOld Highland Park Baptist Church E01Lawrence Garner100% (2)
- CCTV Guidelines - Commission Letter Dated 27.08.2022Document2 pagesCCTV Guidelines - Commission Letter Dated 27.08.2022Sumeet TripathiNo ratings yet
- Bankers ChoiceDocument18 pagesBankers ChoiceArchana ThirunagariNo ratings yet
- Tylenol CrisisDocument2 pagesTylenol CrisisNida SweetNo ratings yet
- Cash Budget Sharpe Corporation S Projected Sales First 8 Month oDocument1 pageCash Budget Sharpe Corporation S Projected Sales First 8 Month oAmit PandeyNo ratings yet
- Elementary Electronics 1968-09-10Document108 pagesElementary Electronics 1968-09-10Jim ToewsNo ratings yet
- CLASS XI (COMPUTER SCIENCE) HALF YEARLY QP Bhopal Region Set-IIDocument4 pagesCLASS XI (COMPUTER SCIENCE) HALF YEARLY QP Bhopal Region Set-IIDeepika AggarwalNo ratings yet
- Bofa Turkish Banks-Back On The RadarDocument15 pagesBofa Turkish Banks-Back On The RadarexperhtmNo ratings yet
- BDRRM Sample Draft EoDocument5 pagesBDRRM Sample Draft EoJezreelJhizelRamosMendozaNo ratings yet
- Saes T 883Document13 pagesSaes T 883luke luckyNo ratings yet
- Written Report in Instructional PlanningDocument6 pagesWritten Report in Instructional PlanningRose Aura HerialesNo ratings yet
- Eureka Math Grade 2 Module 3 Parent Tip Sheet 1Document2 pagesEureka Math Grade 2 Module 3 Parent Tip Sheet 1api-324573119No ratings yet
- Growing Onion Management and Water NeedsDocument25 pagesGrowing Onion Management and Water NeedsKATE NAVAJANo ratings yet
- 30 de Thi Hoc Ky 2 Mon Tieng Anh Lop 9 Co Dap An 2023Document64 pages30 de Thi Hoc Ky 2 Mon Tieng Anh Lop 9 Co Dap An 2023Trần MaiNo ratings yet
- New Compabloc IMCP0002GDocument37 pagesNew Compabloc IMCP0002GAnie Ekpenyong0% (1)
- Knitting in Satellite AntennaDocument4 pagesKnitting in Satellite AntennaBhaswati PandaNo ratings yet
- T Rex PumpDocument4 pagesT Rex PumpWong DaNo ratings yet
- BRENTON TarrantDocument4 pagesBRENTON TarrantSayyidNo ratings yet
- EC105Document14 pagesEC105api-3853441No ratings yet
- ADHD is Awesome: A Guide to (Mostly) Thriving with ADHDFrom EverandADHD is Awesome: A Guide to (Mostly) Thriving with ADHDRating: 5 out of 5 stars5/5 (2)
- Breaking the Habit of Being YourselfFrom EverandBreaking the Habit of Being YourselfRating: 4.5 out of 5 stars4.5/5 (1459)
- Summary of The 4-Hour Body: An Uncommon Guide to Rapid Fat-Loss, Incredible Sex, and Becoming Superhuman by Timothy FerrissFrom EverandSummary of The 4-Hour Body: An Uncommon Guide to Rapid Fat-Loss, Incredible Sex, and Becoming Superhuman by Timothy FerrissRating: 4.5 out of 5 stars4.5/5 (81)
- The Bridesmaid: The addictive psychological thriller that everyone is talking aboutFrom EverandThe Bridesmaid: The addictive psychological thriller that everyone is talking aboutRating: 4 out of 5 stars4/5 (131)
- The Obesity Code: Unlocking the Secrets of Weight LossFrom EverandThe Obesity Code: Unlocking the Secrets of Weight LossRating: 4 out of 5 stars4/5 (6)
- Briefly Perfectly Human: Making an Authentic Life by Getting Real About the EndFrom EverandBriefly Perfectly Human: Making an Authentic Life by Getting Real About the EndNo ratings yet
- Prisoners of Geography: Ten Maps That Explain Everything About the WorldFrom EverandPrisoners of Geography: Ten Maps That Explain Everything About the WorldRating: 4.5 out of 5 stars4.5/5 (1145)
- How to Talk to Anyone: Learn the Secrets of Good Communication and the Little Tricks for Big Success in RelationshipFrom EverandHow to Talk to Anyone: Learn the Secrets of Good Communication and the Little Tricks for Big Success in RelationshipRating: 4.5 out of 5 stars4.5/5 (1135)
- Neville Goddard Master Course to Manifest Your Desires Into Reality Using The Law of Attraction: Learn the Secret to Overcoming Your Current Problems and Limitations, Attaining Your Goals, and Achieving Health, Wealth, Happiness and Success!From EverandNeville Goddard Master Course to Manifest Your Desires Into Reality Using The Law of Attraction: Learn the Secret to Overcoming Your Current Problems and Limitations, Attaining Your Goals, and Achieving Health, Wealth, Happiness and Success!Rating: 5 out of 5 stars5/5 (284)
- Love Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)From EverandLove Life: How to Raise Your Standards, Find Your Person, and Live Happily (No Matter What)No ratings yet
- Gut: the new and revised Sunday Times bestsellerFrom EverandGut: the new and revised Sunday Times bestsellerRating: 4 out of 5 stars4/5 (392)
- No Mud, No Lotus: The Art of Transforming SufferingFrom EverandNo Mud, No Lotus: The Art of Transforming SufferingRating: 5 out of 5 stars5/5 (175)
- Sleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningFrom EverandSleep Stories for Adults: Overcome Insomnia and Find a Peaceful AwakeningRating: 4 out of 5 stars4/5 (3)
- Peaceful Sleep Hypnosis: Meditate & RelaxFrom EverandPeaceful Sleep Hypnosis: Meditate & RelaxRating: 4.5 out of 5 stars4.5/5 (142)
- Outlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisFrom EverandOutlive: The Science and Art of Longevity by Peter Attia: Key Takeaways, Summary & AnalysisRating: 4 out of 5 stars4/5 (1)
- Raising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsFrom EverandRaising Good Humans: A Mindful Guide to Breaking the Cycle of Reactive Parenting and Raising Kind, Confident KidsRating: 4.5 out of 5 stars4.5/5 (170)
- The Happiest Baby on the Block: The New Way to Calm Crying and Help Your Newborn Baby Sleep LongerFrom EverandThe Happiest Baby on the Block: The New Way to Calm Crying and Help Your Newborn Baby Sleep LongerRating: 4.5 out of 5 stars4.5/5 (58)
- The Three Waves of Volunteers & The New EarthFrom EverandThe Three Waves of Volunteers & The New EarthRating: 5 out of 5 stars5/5 (179)
- Love Yourself, Heal Your Life Workbook (Insight Guide)From EverandLove Yourself, Heal Your Life Workbook (Insight Guide)Rating: 5 out of 5 stars5/5 (40)
- The House Mate: A gripping psychological thriller you won't be able to put downFrom EverandThe House Mate: A gripping psychological thriller you won't be able to put downRating: 4 out of 5 stars4/5 (127)
- Summary: I'm Glad My Mom Died: by Jennette McCurdy: Key Takeaways, Summary & AnalysisFrom EverandSummary: I'm Glad My Mom Died: by Jennette McCurdy: Key Takeaways, Summary & AnalysisRating: 4.5 out of 5 stars4.5/5 (2)
- The Other Significant Others: Reimagining Life with Friendship at the CenterFrom EverandThe Other Significant Others: Reimagining Life with Friendship at the CenterRating: 4 out of 5 stars4/5 (1)
- Forever Strong: A New, Science-Based Strategy for Aging WellFrom EverandForever Strong: A New, Science-Based Strategy for Aging WellNo ratings yet
- Summary of The Art of Seduction by Robert GreeneFrom EverandSummary of The Art of Seduction by Robert GreeneRating: 4 out of 5 stars4/5 (46)
- The Secret of the Golden Flower: A Chinese Book Of LifeFrom EverandThe Secret of the Golden Flower: A Chinese Book Of LifeRating: 5 out of 5 stars5/5 (4)