You might also like
- Skraćenice U Kliničkim IspitivanjimaDocument17 pagesSkraćenice U Kliničkim IspitivanjimaNenad RadosavljevićNo ratings yet
- VAT GlossaryDocument1 pageVAT GlossaryNenad RadosavljevićNo ratings yet
- Vokabular Engleskog Jezika Medicinske Struke PDFDocument267 pagesVokabular Engleskog Jezika Medicinske Struke PDFRaša MladenovićNo ratings yet
- Vokabular Engleskog Jezika Medicinske Struke PDFDocument267 pagesVokabular Engleskog Jezika Medicinske Struke PDFRaša MladenovićNo ratings yet
- Bol U Grudima - 2Document41 pagesBol U Grudima - 2Nenad RadosavljevićNo ratings yet
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (895)
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5794)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (399)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (73)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (838)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (588)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (344)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1090)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (120)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- Visual Diagnosis in Emergency Medicine: Arthus ReactionDocument2 pagesVisual Diagnosis in Emergency Medicine: Arthus ReactionKarolina EspalzaNo ratings yet
- Automatic Chemiluminescence Analyzer: Maccura Biotechnology Co., LTDDocument6 pagesAutomatic Chemiluminescence Analyzer: Maccura Biotechnology Co., LTDFranklin soca rojasNo ratings yet
- Principles of Disease Control and PreventionDocument13 pagesPrinciples of Disease Control and PreventionAYO NELSON0% (1)
- MUST To KNOW in Blood Banking 1Document19 pagesMUST To KNOW in Blood Banking 1Aya Virtucio100% (1)
- An Interview Between A Doctor and A Reporter Took Place To Answer The Question of The Mass About The Current Situation and About The Different Types of CovidDocument2 pagesAn Interview Between A Doctor and A Reporter Took Place To Answer The Question of The Mass About The Current Situation and About The Different Types of CovidJuliene Ann B. BolusoNo ratings yet
- 1 s2.0 S0196655316001693 MainDocument3 pages1 s2.0 S0196655316001693 MainSandu AlexandraNo ratings yet
- Autoimmune DiseasesDocument27 pagesAutoimmune DiseasesJudy BegilNo ratings yet
- Jurnal Autoimun PDFDocument7 pagesJurnal Autoimun PDFRara Anglis AninditaNo ratings yet
- Art15 PDFDocument6 pagesArt15 PDFNatalia TexidorNo ratings yet
- Pathophysiology of PneumoniaDocument1 pagePathophysiology of Pneumoniaoxidalaj97% (32)
- Ilovepdf Merged 2 PDFDocument307 pagesIlovepdf Merged 2 PDFAhmed ZidanNo ratings yet
- VIROLOGY Chapter 1Document140 pagesVIROLOGY Chapter 1bilisa gechNo ratings yet
- An Update On Perioperative Anaphylaxis (11!04!2016)Document6 pagesAn Update On Perioperative Anaphylaxis (11!04!2016)Maufer AlNo ratings yet
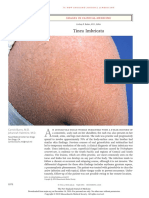
- Tinea ImbricataDocument1 pageTinea ImbricataVirginia AneleyNo ratings yet
- The Silent WarDocument5 pagesThe Silent WarSebastian IguaNo ratings yet
- Chapter 1 (Repaired)Document61 pagesChapter 1 (Repaired)febrinasiskaNo ratings yet
- Kode Icd 10 Bahasa IndonesiaDocument529 pagesKode Icd 10 Bahasa IndonesiaDatin LotengNo ratings yet
- Atherosclerosis PathophysiologyDocument12 pagesAtherosclerosis PathophysiologyHuman ResourcesNo ratings yet
- STING Agonist-Loaded, CD47/PD-L1-targeting Nanoparticles Potentiate Antitumor Immu-Nity and Radiotherapy For GlioblastomaDocument19 pagesSTING Agonist-Loaded, CD47/PD-L1-targeting Nanoparticles Potentiate Antitumor Immu-Nity and Radiotherapy For GlioblastomaAnonymous HUY0yRexYfNo ratings yet
- Community Health Nursing Exam 5: B. Liver CirrhosisDocument7 pagesCommunity Health Nursing Exam 5: B. Liver CirrhosisHannah BuquironNo ratings yet
- Infectious or Communicable DiseasesDocument2 pagesInfectious or Communicable DiseasesДария КоваленкоNo ratings yet
- Pneumonia in Children - Inpatient Treatment - UpToDateDocument23 pagesPneumonia in Children - Inpatient Treatment - UpToDateJunny Yeliitza Wilches GualdronNo ratings yet
- Ca2 Prefinal Exam Pointers To ReviewDocument3 pagesCa2 Prefinal Exam Pointers To ReviewlittlejynxNo ratings yet
- Etiological Potential of Immature Platelet Fraction in Children With ThrombocytopeniaDocument29 pagesEtiological Potential of Immature Platelet Fraction in Children With Thrombocytopeniajhanavi rajeshNo ratings yet
- Presented by Amir Sadaula BVSC & Ah, 8 Sem Rampur Campus Roll No: 01Document19 pagesPresented by Amir Sadaula BVSC & Ah, 8 Sem Rampur Campus Roll No: 01naturalamirNo ratings yet
- ###March2022 - Barangay Monthly Inventory of Vaccinated PopulationDocument2 pages###March2022 - Barangay Monthly Inventory of Vaccinated PopulationBarangay CalaocanNo ratings yet
- HIV & AIDS PathophysiologyDocument3 pagesHIV & AIDS PathophysiologyMarinill Soliman100% (3)
- Reading Comprehension: HIVDocument2 pagesReading Comprehension: HIVFlori75% (8)
- Infeksi Jamur TulangDocument14 pagesInfeksi Jamur TulangNormanPrabowoNo ratings yet
- 3 Wood Flow HematopoieticDocument24 pages3 Wood Flow Hematopoieticalper togayNo ratings yet