You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5814)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1092)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (845)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (540)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (348)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (822)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- Prefixes and Suffixes ListDocument4 pagesPrefixes and Suffixes Listapi-262294171No ratings yet
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
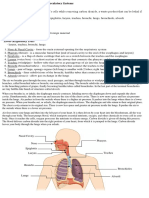
- Delhi Public School, Harni: Subject - Biology (Science) Class - X Chapter-6 Life ProcessesDocument54 pagesDelhi Public School, Harni: Subject - Biology (Science) Class - X Chapter-6 Life ProcessesAnin BertNo ratings yet
- Auriculotherapy For Respiratory DisordersDocument3 pagesAuriculotherapy For Respiratory DisordersCarissa Nichols100% (3)
- Anatomi Dan Fisiologi RespirasiDocument22 pagesAnatomi Dan Fisiologi RespirasinortonaingNo ratings yet
- Lecture (4) Anatomy of Lung & PleuraDocument20 pagesLecture (4) Anatomy of Lung & Pleuracavagnetto maurizioNo ratings yet
- Anatomy of The Thorax MCQDocument7 pagesAnatomy of The Thorax MCQInnocent Clifford Marandu93% (14)
- Anesthesia Review 1000 Questions and Answers To Blast The BASICS and Ace The ADVANCED LWW Wolters KluwerDocument53 pagesAnesthesia Review 1000 Questions and Answers To Blast The BASICS and Ace The ADVANCED LWW Wolters Kluwercrystal.otero76180% (5)
- Clinical Findings in Respiratory SystemDocument1 pageClinical Findings in Respiratory SystemYusri ArifNo ratings yet
- Science Reviewer - Respiratory and Circulatory SystemDocument2 pagesScience Reviewer - Respiratory and Circulatory SystemMaribeth Villanueva100% (2)
- NCM 112 Lecture Notes (Respiratory System and Its Diseases)Document9 pagesNCM 112 Lecture Notes (Respiratory System and Its Diseases)Kyle FernandezNo ratings yet
- Medical Nutrition Therapy For Respiratory DisordersDocument7 pagesMedical Nutrition Therapy For Respiratory DisordersBok MatthewNo ratings yet
- Findings in Respi. AssessmentDocument13 pagesFindings in Respi. AssessmentChristine LeeNo ratings yet
- What Protections Does The Upper Airway Have For The Lower Airway?Document3 pagesWhat Protections Does The Upper Airway Have For The Lower Airway?Paul JacksonNo ratings yet
- NURS1108 Lecture 14 - Respiratory System PART 1 of 2Document62 pagesNURS1108 Lecture 14 - Respiratory System PART 1 of 2Jacia’s SpaceshipNo ratings yet
- Answer Key CicDocument3 pagesAnswer Key Cictiburshoc16No ratings yet
- Lung AnatomyDocument16 pagesLung AnatomywhyseejoshNo ratings yet
- Chest X Ray Techniques and PositioningDocument67 pagesChest X Ray Techniques and PositioningSandeep ChaudharyNo ratings yet
- Master Dentistry2Document275 pagesMaster Dentistry2Isak Shatika0% (1)
- Med Surg Respiratory SystemDocument173 pagesMed Surg Respiratory Systembamfalcon100% (4)
- S 2 Unit 4 Course Book, Work Book Ques and KeysDocument19 pagesS 2 Unit 4 Course Book, Work Book Ques and KeysHein Aung Zin100% (2)
- Pneumothorax (Collapsed Lung)Document34 pagesPneumothorax (Collapsed Lung)james garcia100% (3)
- List of Occupational DiseasesDocument20 pagesList of Occupational DiseasesAbhishek S PillaiNo ratings yet
- Jurnal 1 - Saccus PneumaticusDocument7 pagesJurnal 1 - Saccus PneumaticusZayyan MuafiNo ratings yet
- Medical Terminology PDFDocument48 pagesMedical Terminology PDFtiffylolaNo ratings yet
- Why Do You Need To Breathe? - To LiveDocument5 pagesWhy Do You Need To Breathe? - To LiveJuan DavisNo ratings yet
- The Respiratory SystemDocument26 pagesThe Respiratory SystemdanrcgNo ratings yet
- 11.2 Gas Exchange in Humans Igcse Cie Biology Ext Theory QPDocument5 pages11.2 Gas Exchange in Humans Igcse Cie Biology Ext Theory QPhamnafayyaz1995No ratings yet
- Turpentine TherapyDocument5 pagesTurpentine Therapyvivesur100% (1)
- Imaging Pulmonary Infection, Classic Sign and Pattern PRESENTASIDocument84 pagesImaging Pulmonary Infection, Classic Sign and Pattern PRESENTASIMark Brown100% (1)
- 13.postural DrainageDocument4 pages13.postural DrainageSharon LawrenceNo ratings yet