You might also like
- The Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeFrom EverandThe Subtle Art of Not Giving a F*ck: A Counterintuitive Approach to Living a Good LifeRating: 4 out of 5 stars4/5 (5807)
- The Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreFrom EverandThe Gifts of Imperfection: Let Go of Who You Think You're Supposed to Be and Embrace Who You AreRating: 4 out of 5 stars4/5 (1091)
- Never Split the Difference: Negotiating As If Your Life Depended On ItFrom EverandNever Split the Difference: Negotiating As If Your Life Depended On ItRating: 4.5 out of 5 stars4.5/5 (842)
- Grit: The Power of Passion and PerseveranceFrom EverandGrit: The Power of Passion and PerseveranceRating: 4 out of 5 stars4/5 (590)
- Hidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceFrom EverandHidden Figures: The American Dream and the Untold Story of the Black Women Mathematicians Who Helped Win the Space RaceRating: 4 out of 5 stars4/5 (897)
- Shoe Dog: A Memoir by the Creator of NikeFrom EverandShoe Dog: A Memoir by the Creator of NikeRating: 4.5 out of 5 stars4.5/5 (537)
- The Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersFrom EverandThe Hard Thing About Hard Things: Building a Business When There Are No Easy AnswersRating: 4.5 out of 5 stars4.5/5 (345)
- Elon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureFrom EverandElon Musk: Tesla, SpaceX, and the Quest for a Fantastic FutureRating: 4.5 out of 5 stars4.5/5 (474)
- Her Body and Other Parties: StoriesFrom EverandHer Body and Other Parties: StoriesRating: 4 out of 5 stars4/5 (821)
- The Emperor of All Maladies: A Biography of CancerFrom EverandThe Emperor of All Maladies: A Biography of CancerRating: 4.5 out of 5 stars4.5/5 (271)
- The Sympathizer: A Novel (Pulitzer Prize for Fiction)From EverandThe Sympathizer: A Novel (Pulitzer Prize for Fiction)Rating: 4.5 out of 5 stars4.5/5 (122)
- The Little Book of Hygge: Danish Secrets to Happy LivingFrom EverandThe Little Book of Hygge: Danish Secrets to Happy LivingRating: 3.5 out of 5 stars3.5/5 (401)
- The World Is Flat 3.0: A Brief History of the Twenty-first CenturyFrom EverandThe World Is Flat 3.0: A Brief History of the Twenty-first CenturyRating: 3.5 out of 5 stars3.5/5 (2259)
- The Yellow House: A Memoir (2019 National Book Award Winner)From EverandThe Yellow House: A Memoir (2019 National Book Award Winner)Rating: 4 out of 5 stars4/5 (98)
- Devil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaFrom EverandDevil in the Grove: Thurgood Marshall, the Groveland Boys, and the Dawn of a New AmericaRating: 4.5 out of 5 stars4.5/5 (266)
- A Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryFrom EverandA Heartbreaking Work Of Staggering Genius: A Memoir Based on a True StoryRating: 3.5 out of 5 stars3.5/5 (231)
- Team of Rivals: The Political Genius of Abraham LincolnFrom EverandTeam of Rivals: The Political Genius of Abraham LincolnRating: 4.5 out of 5 stars4.5/5 (234)
- On Fire: The (Burning) Case for a Green New DealFrom EverandOn Fire: The (Burning) Case for a Green New DealRating: 4 out of 5 stars4/5 (74)
- The Unwinding: An Inner History of the New AmericaFrom EverandThe Unwinding: An Inner History of the New AmericaRating: 4 out of 5 stars4/5 (45)
- Ballantyne - The Massachusetts General Hospital Handbook ofDocument229 pagesBallantyne - The Massachusetts General Hospital Handbook ofadyna_popa_79462000850% (2)
- Test 7 & 8 BDSDocument12 pagesTest 7 & 8 BDSrababNo ratings yet
- External & Internal Features of Spinal CordDocument77 pagesExternal & Internal Features of Spinal CordPatricia Hariramani100% (2)
- Pain ManagementDocument17 pagesPain Managementa bNo ratings yet
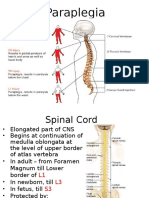
- ParaplegiaDocument51 pagesParaplegiaChuah Wei HongNo ratings yet
- Spinal Cord and Nerve Plexii: by Dr. TDocument20 pagesSpinal Cord and Nerve Plexii: by Dr. TthwisemanNo ratings yet
- 1184bns 1Document14 pages1184bns 1Daniela GanchasNo ratings yet
- An Overview of The Physiology of Pain For The VeterinarianDocument5 pagesAn Overview of The Physiology of Pain For The VeterinarianMr. questionNo ratings yet
- Anatomy of Spinal CordDocument41 pagesAnatomy of Spinal CordI Wayan Aryanta PutraNo ratings yet
- 2 NeuroanatomyDocument60 pages2 NeuroanatomyMeirina KhairatNo ratings yet
- 1.5 DermatomesDocument2 pages1.5 DermatomesAngela CaguitlaNo ratings yet
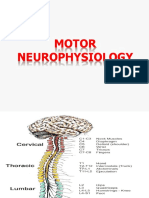
- Genral Motor Function, SC and Muscle SpindlesDocument41 pagesGenral Motor Function, SC and Muscle SpindlesnoufNo ratings yet
- Sem-3 Physiology of PainDocument69 pagesSem-3 Physiology of PainJyoti Pol SherkhaneNo ratings yet
- IAPHD Hand Book On Palliative Care PDFDocument37 pagesIAPHD Hand Book On Palliative Care PDFMohammed ArshadNo ratings yet
- Spinal Cord ExternalDocument33 pagesSpinal Cord ExternalCosplay AccountNo ratings yet
- Kyyrönen IidaDocument114 pagesKyyrönen IidaNopri JayaNo ratings yet
- The Descending Tracts of The Spinal CordDocument2 pagesThe Descending Tracts of The Spinal Cordalmastar officeNo ratings yet
- Sensitasi SentralDocument13 pagesSensitasi SentralRay SirvelNo ratings yet
- Applied Anatomy of Spinal CordDocument34 pagesApplied Anatomy of Spinal CordSuhana PathanNo ratings yet
- Efficacy of Muscle Energy Technique Versus MyofascialDocument7 pagesEfficacy of Muscle Energy Technique Versus Myofascialmuhammad yaminNo ratings yet
- Ascending Tracts Neuro AnatomyDocument2 pagesAscending Tracts Neuro AnatomyrachellesliedeleonNo ratings yet
- Articulo Ingles QuemadurasDocument17 pagesArticulo Ingles QuemadurasAndrea RiveraNo ratings yet
- Acute Service Pain Handbook 2011Document116 pagesAcute Service Pain Handbook 2011Iskandar414No ratings yet
- Pain and Pain Gate Control MechanismDocument17 pagesPain and Pain Gate Control MechanismKapil LakhwaraNo ratings yet
- 2015 Is Neuroplasticity in The Central Nervous SystemDocument14 pages2015 Is Neuroplasticity in The Central Nervous SystemcinNo ratings yet
- Neuroanatomy NotesDocument51 pagesNeuroanatomy NotesAditi MangalNo ratings yet
- Notes On CNS Physiology by Medical Study CenterDocument99 pagesNotes On CNS Physiology by Medical Study CenterHifza Faqeer KambohNo ratings yet